
세균
- 형질전환체 만들기 : 플라스미드 DNA를 세균 안에 넣기
-
CaCl2 : 플라스미드가 세포에 달라 붙는다.
-
42℃ 열충격 : 세포안으로 들어간다.
-
Phenotypic expression(표현형 발현)할 시간 주기 : 37℃, 1 hour (그렇지 않으면 항생제 내성유전자가 들어갔어도 사멸 할 수 있음)
-
- 재조합체 선별하기 : Selection marker 이용하기
- Insertional inactivation : 유전자가 성공적으로 ligation 됐냐
- ampicilin resistance
- tetracyclin resistance ← BamH1로 자른다. 여기에 새 유전자 삽입
- Selectable marker : 비형질전환체가 가지고 있지 않은 새로운 특징을 형질전환된 세포에 제공하는 유전자를 말한다.
- Two step selection (ampR, tetR 포함하는 벡터)
- ampicilin 배지에 깐다 : normal vector 뒤짐
- tetracyclin 배지에 깐다 : recombinant vector 뒤짐
- original로 돌아가서 recombinant vector 죽은 자리의 애들 건진다.
- One step selection (ampR, lacZ' 포함하는 벡터, lacZ : lactose를 분해)
- ampicilin 처리한다 : normal colony 뒤짐
- X-gal, IPTG 처리한다
- X-gal ≒ lactose, 가수분해 되면 파란색 띤다 : Blue colony : 재조합 안된 vector가 들어간 colony
- lacZ 유전자가 발현되지 않는 (lactose 분해 안 된) White color colony : 재조합 된 vector가 들어간 colony
- Insertional inactivation : 유전자가 성공적으로 ligation 됐냐
바이러스
자기가 알아서 recombinant DNA 넣는다.
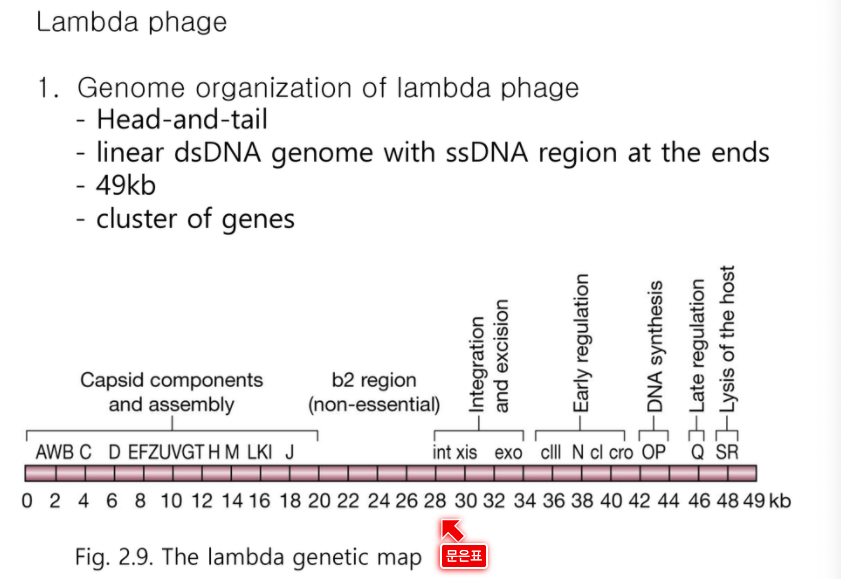
- 람다 페이즈
- Single-strain packaging System
- Two-strain packaging System
- λ packaging System
- 37~52 kb의 DNA분자만을 삽입시킬 수 있다.
- 37 kb보다 작은 어떤 것도 packaging 될 수 없다.
- λ DNA의 거대한 부위를 결손시킴으로써 37 kb보다 작은 λ vector를 제조한다.
- 이들은 extra DNA가 삽입되어 전체 genome size가 37 kb이상이 되는 경우에만 packaging 될 것이기 때문에 recombinant phage만 복제될 수 있다.
- clear plaque : recombinant
- cos site만큼 자르면서 packaging 된다.
- M13 페이즈
동물세포, 식물세포
- Fusion with liposome
- Transformation of plant Protoplast
- 물리적으로 DNA 넣는법
- Microinjection
- 입자총 기법
- Gene gun
DNA를 세포에서 꺼내면 벡터를 자르고 집어 넣는 것이다.
집어 넣는 것을 ligation이라고 한다.
1. DNA 도입하기.
2. 확인하기
3. 운반체로 사용되는 파지 DNA를 도입하기
4. phage 재조합
5. 식물,동물 세포
5.1 형질전환체 만들기 : 세포에 DNA 도입 (transformation)
플라스미드

여기에 도입 한다음에 기르면 엄청난 재조합 DNA를 얻을 수 있다.
삽입한 유전자의 산물을 얻을 수 있다.
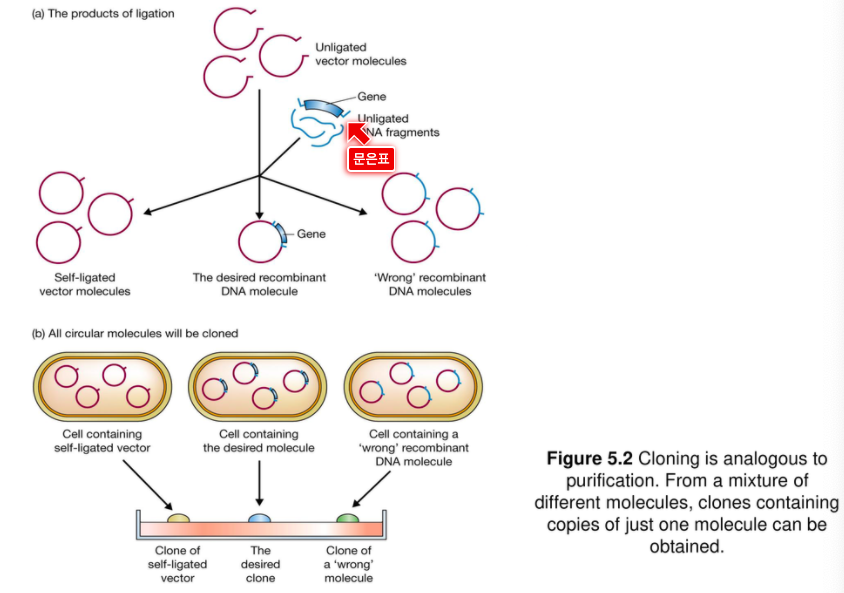
5. 생세포에서의 DNA 도입(형질전환 : Transformation)
재조합 DNA 제조 후 다음 단계는 생세포(보통 세균)안으로 재조합 DNA를 도입시키는 것이며 그 세포는 클론을 생산하기 위해 성장, 분열하게 된다. 엄밀히 말해서, 클로닝이란 재조합 DNA제조 자체를 일컫는 것이 아니라 그 이후 과정만을 말한다.
* 유전자 클로닝의 목적
1) 소량의 DNA로부터 많은 양의 DNA를 얻기 위해
-소량의 DNA로부터 대량의 재조합 DNA를 생산함으로써 유전자 구조와 발현의 연구를 위해 필요한 대량의 DNA 제공
2) 특정 유전자의 산물만을 순수하게 대량으로 얻기 위해
-정제와 유사하게 특정유전자를 가진 재조합체를 선별 가능
일단 순수분리 안하고 섞여있는 상태에서 집어넣고 재조합된 것만 뺀다.
라이브러리를 만든다.
ligation 효율이 낮다
다 섞여있는 상황에서 숙주세포에 싹 다 집어 넣는다.
(b)에서 나타난 3가지 보다 훨씬 더 많은 건 아무것도 안들어간 클론이다.
운반체가 들어간 애들을 먼저 살릴 수 있다.
운반체는 selection marker가 있기 때문에, 특정 항생제를 넣어주면 아무것도 안들어간 꽝인 숙주세포를 버릴 수 있다.
1시간 정도 시간을 주고 selection media 위에 깔아야 한다.

이건 플라스미드 재조합 DNA를 어떻게 넣느냐 이다.
차가운 CaCl2 로 몇 번 씻어주면 칼슘 이온들이 플라스미드가 세포주에 달라붙게 해준다.
그리고 42℃로 열 충격을 주면 flexible하게 들어오게 된다.
DNA가 유전물질이다 라는 증명 실험에서 폐렴균가지고 한 실험을 기억해보아라
S타입과 R타입
독성의 폐렴균
자연계의 박테리아에서 외부 DNA를 받아들이기에 유능한 세포를 처리해줘야 한다. : CaCl2
이것만 가지고 안되기 때문에 42℃에서 열충격을 준다.
이래도 10^6 중 하나만 들어간다.

이렇게 집어 넣은 다음에 바로 선별 배지에 깔면 또 안된다.
도입한 다음에 바로 깔면 안된다

Pheonotypic expression 할 시간을 줘야한다.
표현형 발현할 시간을 줘야한다.
안그러면 항생제 내성유전자가 들어갔어도 죽을 수가 있다. ★★★★★★★
5.2 재조합체 선별하기

플라스미드의 선별 마커가 있다.
들어간 애만 살리기 위해서 항생제 내성 유전자가 있다.
그러나 그 이전에
재조합된 애들을 만들기 너무 힘들기 때문에
대부분이 삽입이 안돼있어서 이걸 또 구분할 수 있어야한다.
우리 유전자를 어디에다 넣을 것이냐?
어떤 유전자A를 잘라서 넣어야한다.
대부분은 그냥 닫혀버린다.
그러다 어쩌다 내 유전자가 들어간게 있을 것이다.
ligation이 성공적으로 됐으면 잘라낸 이 유전자A는 망가진다.
선별마커를 두개가 있어야 한다.
이 플라스미드가 들어갔냐 안들어갔냐
유전자가 성공적으로 ligation이 됐냐. 유전자A가 망가졌느냐 : insertional inactivation
Two Step Selection

Ampicillin resistance
BamH1
(a)는 엠피실린 항생제, 테트라사이클 항생제에서 살아남는다.
(b)는 엠피실린 항생제에선 살고, 테트라사이클 항생제에서는 죽는다.
처음부터 두개를 넣지 않고 Two step selection 을 한다

선별배지에 깐 다음에 항생제 내성 유전자 두개가 다 있는 배지를 쓰지 않고
처음에 엠피실린만 있는 배지에다가 깐다.
그러면 (a) normal vector도 살고 (b) recombinant vector 도 산다.
아무것도 안 들어간 애들은 죽는다.
스탬프를 찍으면 형질전환체 일부가 올라간다.
위에서 자라는데 아래에서 안자라는 애들이 찐이다.
그래서 오리지날로 돌아가서 안자라는 애들을 건져야 한다.
이게 Two step selection이다
One step selection
★★★★★★★

엠피실린 내성유전자 남겨놓는다.
두번째 선별 마커는 락토오스 대장균
lacZ 를 쓰면 알 수 있다.
BamH1은 망가진다.
블루가 아니고 화이트 칼라가 된다. lacZ 가 망가지니까 blue를 못낸다.
lacZ는 뭐고 lacZ ' 프라임은 뭐냐?
프라임은 lacZ의 일부이다.

lacZ 의 일부만 넣는다.
lacZ 라는 유전자는 잘라서 두개를 따로따로 세포에 넣어도 가까이에서 만들어지는게 합해져서 기능을 할 수 있다는 걸 알아냈다.
(a)는 엠피실린에 sensitive하고 lacZ 가 온전하지가 않기 때문에, 프라임이 없기 때문에 blue 칼라를 못 만든다.
(a)의 오른쪽 것은 lacZ 유전자가 너무 크기 때문에 온전한 lacZ를 플라스미드 마커를 넣지 않고 일부분만 쓴다.
pUC8의 lacZ 프라임 부분을 열어서 우리 유전자를 넣는다.
배지에 X-gal을 넣어준다.
염색제인 X-gal이 있는 평판배지에서 세포들이 자라나고
접합 성공 / 실패 여부에 따라 콜로니가 다른 색을 띄게 된다.
콜로니의 색 차이가 이 기술의 핵심이다.
접합 성공 : White ●
접함 실패 : Blue ●
배지에다가 X-gal IPTG 엠피실린 넣어준다.
white color가 내가 원하는 배지이다.
다 기억해라
Blue-white screening 은 amp 와 lacZ를 포함하고 있는 벡터인 pUC를 이용한다.
여기에서 amp 는 항생제인 ampicilin에 저항성을 갖는 유전자이고,
lacZ 는 β-galactosidase 에 관한 유전자이며
β-galactosidase는 lactose 를 galactose와 glucose로 가수분해하는 효소이다.
형질 변환이 제대로 되지 않은 균주의 군락은
X-gal이 β-galactosidase 활성에 의해 가수분해 되면서 파란색을 띠게 된다.
왜냐하면 X-gal이 lactose의 유사체(analogue)여셔, β-galactosidase 에 의해 lactose로 인식되어 분해되는 것이다.
β-galactosidase 를 발현시킬 수 있는 lacZ는 pUC의 다복제구역(multiple cloning site, MSC)에 위치하고 있어서
만약 외래 DNA가 MSC 안으로 삽입되게 되면 lacZ는 비활성되어 β-galactosidase 가 발현되지 않는다.
따라서 원하는 DNA가 삽입된 재조합 플라스미드 DNA가 제대로 도입된 형질전환세포는
lacZ 가 비활성되기 때문에 X-gal 분해능력이 없어서 배지에서 하얀색을 띠는 콜로니를 형성하는 것이다.

5.3 Introduction of phage DNA into bacterial cells
람다 페이즈

람다가 DNA를 쭉 집어 넣는다.
끝에 단일가닥 점착성말단으로 원형이 된다.
recombination해서 통합된다. 자연계에 이런게 있다.
그런데 조건을 주면 람다가 엄청나게 튀어 나온다 : lytic cycle
C1 gene이 망가지면
M13 페이즈
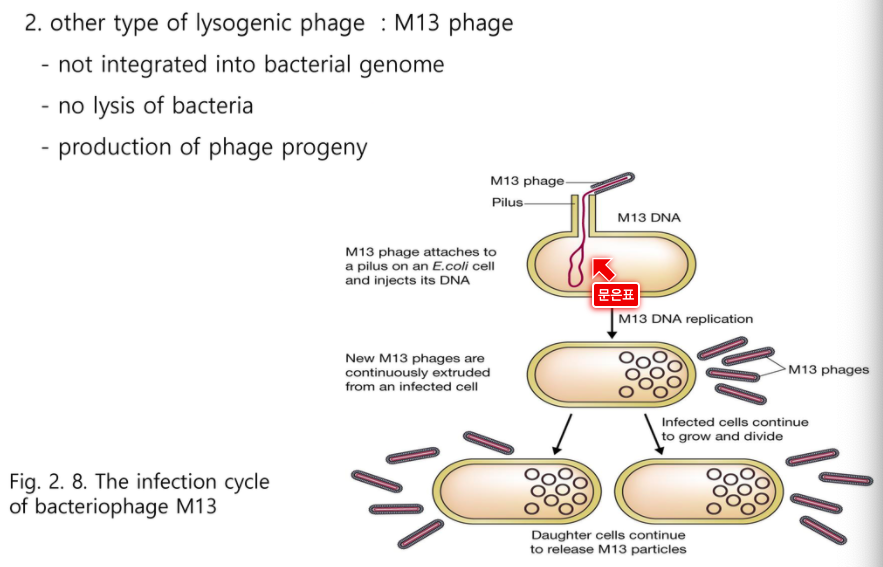
이 단일 가닥은 자기 DNA를 만들려면 이것에 상보적인 가닥이 있어야한다.
여러개가 복제되면 껍데기가 만들어지고
세포를 깨고 나오는게 아니라 분비의 형태로 나온다.
M13은 숙주를 깨지 않는다. 분비한다.
복습해 준것

1) Single-strain system
cos site에 돌연변이를 가지고 있기 때문에 λ concatamer를 정상적으로 절단하는 endonuclease에 의해 인식되지 못하는 defective λ phage를 이용한다. 이 defective phage는 packaging에 필요한 모든 단백질을 합성함에도 불구하고 복제할 수 없다. 이 단백질들은 세균안에 축적되어 있으므로 숙주세포를 파괴시키고 미리 준비한 정상적인 cos site를 포함하고 있는 재조합 DNA와 혼합시키면 재조합 DNA가 capsid내로 들어감으로써 완전한 λ phage가 된다.
① cos site사이에 foreign DNA를 삽입
② λ-defective strain 사용 : 이 strain은 cos site가 변형되어 concatamer가 절단되지 않는 λ로 structural protein은 정상적으로 만들어지지만 DNA가 들어가지 않는다(크기가 너무 크기 때문)
③ ②가 감염된 host cell을 파괴시키고 미리 제조하여 준비한 ①을 첨가하면 정상적인 cos site가 절단되어 foreign DNA를 포함한 recombinant λ DNA가 ②의 head(capsid)에 들어감으로써 in vitro packaging시켜 완전한 phage λ를 제조
2) Two-strain system
서로 다른 phage protein의 유전자(gene D, E)에 각각 돌연변이를 가진 두 종류의 defective λ phage를 각각 다른 숙주세포에서 배양시킨다. 완전한 capsid가 만들어질 수는 없지만 다른(나머지) 모든 coat protein들은 축적된다. 두 숙주세포를 파괴 후 재조합 DNA와 혼합시키면 in vitro packaging이 일어난다.
①서로 다른 phage protein gene에 mutation이 일어난 2종류의 defective λ strain을 이용
②각각 다른 host에서 증식시킨 후, host cell을 파괴, 혼합하고 recombinant λ DNA를 첨가하여 in vitro packaging시키는 방법
람다 genome을 꺼내서 우리 유전자를 삽입 했다.
플라스미드는 꺼내기 쉽고 조작하기 쉽고 별도의 처리없이 섞어서 해서 편하다.
바이러스의 장점은 복잡한 것이다. 플라스미드보다 생물에 가깝기 때문.
그런데 바이러스의 운반체의 가장 큰 장점은 자기가 알아서 재조합DNA를 집어넣는다는 것이다.
형질전환이 어렵지 않다는 것이다.
지가 알아서 다하기 때문.
람다가 자기 DNA를 쭉 주입하면 껍데기 단백질들 막 만든다.
그러면 알아서 껍데기가 조립이 되면서
(c)
람다 껍데기를 벗겨내고 람다 DNA를 얻는다.
근데 이걸 넣어도 안된다.
껍데기를 가지고 있어야 한다.
껍데기 단백질을 주워 모아야한다.
(a)에서 껍데기 단백질 모은다.
5.3.1 Phage infection is visualized as plaques on an agar medium
* plaque : bacterial lawn상에, phage에 의한 cell lysis에 의해 형성된 clear zone.
-lytic plaque : 모든 bacteria가 lysis되고 높은 phage titre를 얻을 수 있다.
-M13 plaque(tubid plaque) : lysogenic phage에 의해 형성되며, plaque에는 slow-growing bacteria와 M13 phage가 존재

결과를 보면 람다를 만들어서 박테리아에 섞었다.
그런 다음에 박테리아 대장균을 넣어서 증식하고 고체 배지 위에다가 깔면
재조합 람다들이 계속 깨고 나오고 깨고 나오고 해서 clear zone이 생성된다.
여기에는 다량의 재조합 람다와 죽은 박테리아가 있다.
M13은 반투명하게 생긴다. 그림과 살짝다름
느슨하게 분열한다.
M13가 모든걸 가져가서 자기가 증식하는데 써서 대장균이 빽빽하지 못하고 반투명한 플라크가 생긴다.
5.4 Identification of recombinant phages

lacZ 는 (a) 에서 여기서 항생제 내성은 넣어주지 않는다.
(b) cl gene을 짤라서 넣어주면 통합이 안되고 마구마구 증식해서 클리어존이 된다.
람다가 끼어들어가지 않는 곳보다 늦게 자란다. : Turbid 플라크
(c) Spi 라는 것은 p라는 것이 p2라는 대장균의 바이러스이다
P2바이러스는 genome이 삽입되어 있다.
이렇게 삽입돼 있을 때 페이즈라고 안하고 프로페이즈 라고 부른다.
이런 대장균을 숙주로 쓴다.
spi 가 (+) 면 들어가지 못하고
spi 가 (-) 면 들어갈 수 있다.
잘라서 넣게되면 재조합체니까. 잘라져서 망가져야면 형질전환이 된다.
(d)는 대용량으로 만들 때 중요하니까 특별히 잘 기억을 해둬라.
세포 안에 들어가는데 람다 유전자를 꺼낸 다음에 (그대로 쓰기힘들다 커서)
줄여서 그 차이만큼 우리 유전자를 집어 넣는다.
자기 원래 유전자보다 작다. 람다가.
적당한 크기 우리유전자가 들어가면 원래 자기 사이즈와 같아진다.
우리가 원하는 것은 correct size 보다 살짝 큰 부분만 잘라서 증식하게 해야한다.
λ packaging system은 37~52 kb의 DNA분자만을 삽입시킬 수 있다.
37 kb보다 작은 어떤 것도 packaging될 수 없다.
λ DNA의 거대한 부위를 결손시킴으로써 37 kb보다 작은 λ vector를 제조한다.
이들은 extra DNA가 삽입되어 전체 genome size가 37 kb이상이 되는 경우에만 packaging 될 것이기 때문에 recombinant phage만 복제될 수 있다.
불필요한 부분자르고
람다가 자기자신을 인정하는 것은 사이즈이다.
우리유전자를 삽입해서 원래 크기가 되면 패키징을 한다.
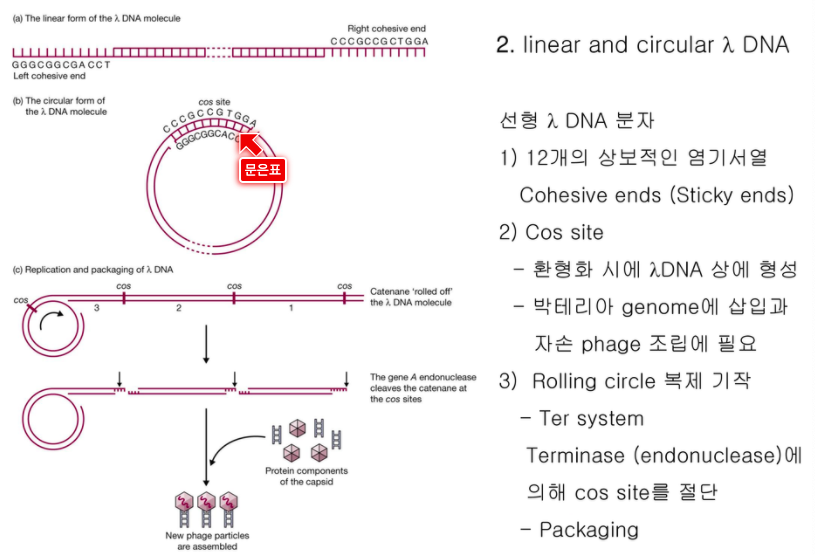
람다가 복제되면 줄줄이 사탕으로 복제된다.
termianse라는 걸로 cos site를 잘라서 점착성말단의 한 단위 지놈으로 자른다.
자식으로 인정되면 람다페이즈를 만든다.
재조합체만 원래사이즈로 인식해서. (플라스미드처럼 두번째 마커가 필요 없다)
아까 (d) 잘 기억해둬라
동물세포 식물세포

박테리아보다 동물에 넣기가 더 쉽다.
재조합 DNA가 침전되고 안으로 들어간다.
2) 리포좀 을 통한것
지질 2중층
3) 세포벽을 제거하면 안에 세포막만 남는 프로토플라스트가 되고 이 때 넣는다.
그래도 우리가 원하는건 재조합 식물이니까 적절하게 배지에 넣어서 키운다.
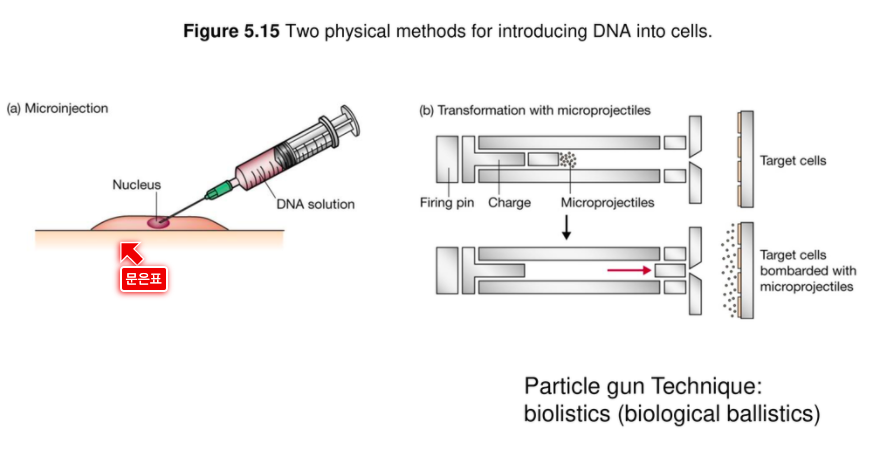
미세주사기로 핵 정조준해서 안에 집어 넣기.
(b) 입자 총 기법
미세 탄환에다가 재조합체 DNA를 코팅한 다음에 총알에 미세 탄환을 집어 넣는다.
미세탄환이 튀어나가면서 Target cell 안에 들어간다.

이게 미세탄환 : DNA 코팅해서

응용한게 gene gun
입자 총 기법은 식물에서 굉장히 많이 쓰인다.
캘러스에 입자총을 쏴서 캘러스를 배지에 넣으면 더 잘된다.
'🧬 Bio > 유전공학' 카테고리의 다른 글
| 유전공학 7장 - 대장균 이외의 다른 생물체 용도의 클로닝 벡터 (0) | 2020.04.09 |
|---|---|
| 유전공학 6장 - 대장균(E. coli) 용도의 클로닝 벡터 (0) | 2020.04.06 |
| 유전공학 4장 - Manipulation of purified DNA (0) | 2020.03.30 |
| 유전공학 4장 - Electrophoresis (0) | 2020.03.26 |
| 유전공학 4장 - DNA manipulation (0) | 2020.03.26 |