


Klenow fragment
Taq DNA polymerase
Reverse transcriptase 이때의 주형은 RNA이다.


이중나선에서 DNA의 방향성을 기억해야한다.
5' 3'
3' 5'
많은 효소들이 ligase가 붙일 때 효율이 좋다 sticky end가. 점착성 말단
단일가닥 돌출부위가 나오게 잘라진 = sticky end

제한효소를 자르고 나서 원하는 부분을 골라야 한다.
제한효소로 DNA를 잘랐을 때 여러가지 조각이 나온다.
여러가지 조각이 섞여 있을 때 특정 조각을 분리해내는 방법 : 전기영동
DNA는 pH 7 정도에서 인산기에서 H+가 해리가 되기 때문
DNA는 -극의 반발력과 +극의 인력 때문에 +극 쪽으로 이동하게 된다.
DNA의 전하량을 보면 염기하나 늘어날 때 마다 음전하가 늘어나기 때문에 비율은 일정하다.
그래서 버퍼속에서 움직이게 하지 않고 고체판(젤)에서 움직이게 한다.
아가로스젤에 공기구멍이 생기면서 그 사이사이로 이동한다.
그래서 작은 DNA가 더 빨리 내려가고 큰 DNA가 더 늦게 내려간다.
분자량에 의해서 속도가 달라지는 것은 아가로스 젤에 구멍을 통과할 때 얼마나 제재를 받느냐, 마찰이 생기느냐에 결정된다.

이 DNA는 맨눈으로 볼 수 없고 가시화를 시켜야 한다.
UV 판 위에 올려놓고 보는 것이다.
염색 해주기 <- EtBr
그러면 밝은 오렌지색 빛을 볼 수 있다.
EtBr의 민감도는 10ng이상일 때만 작동한다.
그래서 더 작으면 동위원소로 하던가 형광표지법을 쓴다.

구조는 나중에 보면 차이가 있다.
구멍을 통과해야 하기 때문에 분자량이 중요하다.
아가로스 젤 & 폴리아크릴아마이드 젤
resolution에서 차이가 있다. 언제쓰는지 알아야 한다.
1) 아가로스젤
1 base pair 차이를 보는게 아니라 수십~수백 bp 차이를 볼 때 쓴다.
2) 폴리아크릴아마이드 젤
1 base pair 차이가 나는 것도 분리할 수 있다.

퍼센트가 낮아지면 구멍사이즈가 커진다.
퍼센트가 높아지면 구멍사이즈가 작아진다.
그래서 작은 DNA는 구멍사이즈가 작을 때 큰 DNA보다 잘 내려온다
작은 DNA 볼 때 : 높은 퍼센티지
큰 DNA 볼 때 : 작은 퍼센티지

대략 분자량에 반비례를 한다.
분자량의 로그를 씌운 값에 반비례한다.
방사선 동위원소법 생략한다.
제한효소 지도

이종수 교수님이 설명 하셨음
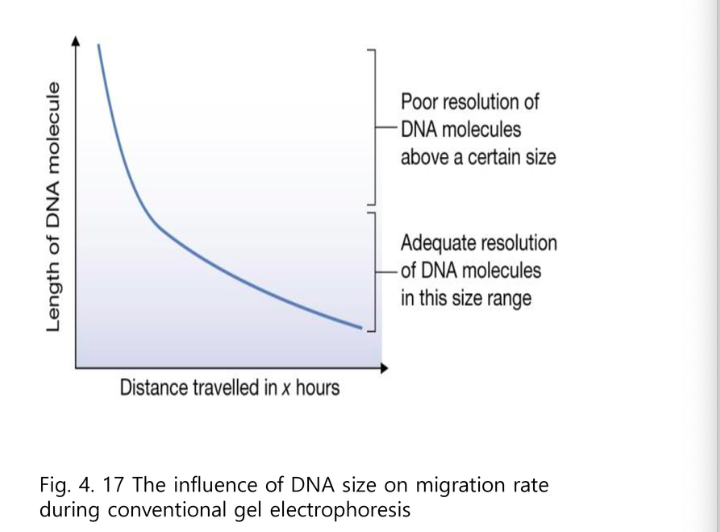
유의할 것 :
반비례해서 이렇게 나오지만
구간의 해상력에 차이가 있다.

1번 구간은 로그에 따라 반비례하는 구간이다
2번 구간은 별 차이가 없다.
굉장히 큰 DNA는 분리하기가 굉장히 어렵다는 것이다.
작은 DNA이기 때문에 분자량이 조금만 차이가 나도 차이가 크다.
빨리가는 애는 이미 빠져 나가버려서 빨리 나간 쪽 부분은 분리를 하기가 힘들다.
퍼센티지를 낮추게 되면 젤이 물컹거려서 굳지가 않는다.

이걸 해결하기 위해서 나온 방법 : PFGE
다 잊어버리고 PFGE로만 기억해라.
지금 까지의 전기영동은 - 극과 +극 한개로 fix되어 있었지만,
PFGE에서는 이 전기장이 교대로 바뀐다.
이 특정 경우에는 맨처음 전기장이



각각의 방향으로 움직이려고 하지만 결국 방향을 정하고 내려간다.
각 전기장에 대해서 DNA가 방향을 정한다. orientation
convention한 젤에서는 아주 큰 DNA나 아주 작은 DNA는 잘 안보인다.
하지만 이건 이틀걸려서 하면 보인다.

자르는 칼과 붙이는 풀이 필요하다
칼 = 제한효소, 풀 = 라이게이스
라이게이스 : 기존에 있는 DNA를 붙인다
물빠지고 인산에스테르결합 형성

blunt end 보다 sticky end가 효율이 훨씬 높다. ****
단일 가닥 부위가 서로 상보적이기 때문에 이미 수소결합이 맞춰져 있어서 ligation이 훨씬 쉽다.

BamH1으로 우리 DNA를 자르고 싶어도 DNA에 없으면 못자른다.
이걸 해결할 방법이 있다.
linker
adaptor
homopolymer
DNA 끝에 linker 를 붙인다.
linker는 엄청나게 붙는다.
그런다음 BamH1으로 자르면 마지막것만 붙게 되고 우리 DNA는 sticky end를 갖게 된다.
우리 유전자 내부에 없는 제한효소 가지를 가진 linker를 써야한다.
이게 쉽지 않기 때문에 adaptor를 쓴다.
두개가 연결

adaptor 끝을 독특하게 한다.

두 조각을 붙이려면 3' 에는 OH 5'에는 인산기가 있어야 한다.
그래야 ligase가 붙일 수 있다.


3번째 방법 : homopolymer
DNA modifying enzyme에 속한다.
우리 DNA가
dCTP만 붙인다.
우리 DNA는 3' DNA만 붙이면 이거를 집어넣으러면 운반체에는 GGGG를 붙여야 한다.
엄청나게 많은가닥을 가지고 CCC를 붙이기 때문에 벡터나 모든 가닥에 붙어있진 않다.

자연상 DNA는 super helix를 갖기 때문에. topoisomerase는 유전공학에서는 잘 쓰지 않았다.
최근에 각광받는다.
헬리카아제가 twist를 풀면 사실은 푸는게 아니라 앞에 꼬임을 쌓아 놓는다.
이게 초나선으로 더 꼬이게 된다.
topoisomerase 중 type1 은 한가닥만 자른다. Nick이 생기게 해서
뒤로 보내서 T값을 감소하게 한다.
이 가닥을 뒤쪽으로 붙이면 풀어지게 된다.

토포아이소머레이슨는 DNA를 자르기도하고 붙이기도 한다. 중요하다.
topo는 DNA를 자르고 이탈되는게 아니라 끝에 붙어있다.
뒤에 방향으로 가서 다시 연결해야 되기 때문에.
벡터를 열면 토포가 붙어있다.
5' 3' blunt end는 효율이 낮다고 했다.
5'의 인산기를 제거한다. Alkaline phosphatase
이미 얘를 붙잡고 있기 때문에 붙인다.
인산에스테르 결합을 시키진 못하지만 쉽게 붙여서 각광받는다.

Vaccinia topoisomerase
type1 이고 CCCTT를 자르고
자른 끝에 T가 붙어있는다. ligation을 쉽게 하려고

끝에 T를 남긴다, 붙인다.
증폭을 했을 때
특별히 PCR product(taq ) 을 유용하게 한다.
제한효소 겸 ligase 역할을 하기 때문에 중요하다.
'🧬 Bio > 유전공학' 카테고리의 다른 글
| 유전공학 6장 - 대장균(E. coli) 용도의 클로닝 벡터 (0) | 2020.04.06 |
|---|---|
| 유전공학 5장 - Introduction of DNA into living cells (0) | 2020.04.02 |
| 유전공학 4장 - Electrophoresis (0) | 2020.03.26 |
| 유전공학 4장 - DNA manipulation (0) | 2020.03.26 |
| 유전공학 3장 (2) (0) | 2020.03.23 |