미생물학 1장
휘태커 5계 분류 체계
five kindom = 5계 분류체계
- 모네라 (원핵세포)
- 원생생물
- 진균
- 동물
- 식물
Woese 3영역 분류체계를 보아라
이렇게 계통수 확립을 할 수 있도록 크게 3영역으로 나눴다. 바이러스는 3영역 분류체계에 들어가지 않는다.
- 세균
- 고세균
- 진핵생물
라이보자임
- 가장 최초에 나타나서 복제하고, 합성하기위해 RNA를 사용했다.
- 효소의 기능을 가진 RNA
남세균 : 최초로 산소를 만들어낸 생물
계통분석 : rRNA의 염기서열을 이용한다.
내부공생설 증거
- 세균의 유전체 서열이 비슷하다.
- 막이 두개이다.
- 자체 DNA
- 수소가설 : 산소가 없었을 당시 중요한 가설
미생물학의 기원
- 레벤후크 : 현미경 최초 발견, 자기 구강세균 발견
- 파스퇴르 : 자연발생설 (백조목 플라스크 실험), 백신 만듦
- 코흐 : 미생물 병인론 (탄저균에 의해 탄저병 일어난다는 것을 증명), 영양배지 개발
- 리스터 : 페놀이라는 소독제 발견
- 바이아링크 : 최초의 바이러스 발견
미생물학 2장 (1) - 세균의 세포구조
- 1 나노미터 = 10 옹스트롬
- 1 옹스트롬 = 0.1 나노미터
- 원형질막 (100 옹스트롬)
- 인지질 : 양친매성
- 호파노이드 : 세균의 스테로이드 : 막안정성 유지
- 세균의 호흡이 원형질막에서 일어난다.
- 핌브리아 : binding에 중용한 역할
- pili : DNA,염색체 주고 받음
- 촉진확산 : 에너지가 필요없는 수송
- 통로단백질 (포화 X)
- 운반단백질 (포화 O)
- 수동확산 : 막을 통해 이동
- 1차 능동수송
- 기질단백질에 용질이 결합
- ATP 가수분해
- ABC transporter를 통해 이동한다
- ABC 수송체에 한 분자가 통과하려면? ATP 2분자 필요
- 2차 능동수송
- 두가지 물질이 동시에 수송된다.
- 공동수송체(같은 방향) & 역수송체(다른 방향)
- 작용기 전달
- 물질이 세포 안으로 들어오면서 화학적 변형이 일어나는것
- 작용기가 전달이 되면서 수송이 된다.
- ex) 만니톨은 IIC 통로를 통해 들어오면 인산화가 된다 -> 만니톨-1-P는 바로 대사에 참여
- 그람양성 : 펩티도글리칸 - 주변세포질 - 원형질막 - 원형질
- 그람음성 : 외막 - 주변세포질 - 펩티도글리칸 - 주변세포질 - 원형질막 - 원형질
- 외막이 없다면 그람 양성이다.
- 펩티도글리칸의 구조의 특징 : 망상구조(그물구조)
미생물학 2장 (2) - 세균의 세포벽, 외피, 리보솜, 편모
- 테이코산 [그람양성]
- 그람 양성균에만 존재한다
- 세포벽을 견고하게 한다.
- 외피의 음성화(-)는 숙주세표면에 잘 결합하게 해준다.
- 지질다당체 [그람음성 외막]
- 항원 역할하는 O항원 존재
- 구조 : 지질A - 중심다당체 - O항원
- 포린단백질 [그람음성 세포벽]
- 외막의 투과성을 높여준다.
- 단당류의 출입을 도와준다.
- 페니실린 : 새로 만들어지는 세포벽의 합성을 억제시킨다.
- 페니실린 처리 : 그람양성 -> 프로토플라스트
- 페니실린 처리 : 그람음성 -> 스페로플라스트
- 라이소자임 : NAM-NAG 자른다. 세포벽 자른다.
- 외피 : 협막, 점막 존재
- 협막은 조직화 : 세균의 부착 ****
- 점막은 비조직화 : 세균의 이동
- Glycocalyx는 협막과 점막을 모두 포함하는 개념
- 세포골격
- FtsZ (미세소관) : 격벽을 형성한다
- CreS (중간섬유) : 곡선 모양 형성
- MreB (미세섬유) : 막대 모양 유지, 세균 모양 유지
- 카복시솜 : 이산화탄소 저장
- 세균 리보솜
- 70S 리보솜 = 50S 소단위 + 30S 소단위
- 30S 소단위의 16s rRNA로 서열 분석
- 직경 = 200 옹스트롬
- 그람음성세균 편모
- 4개의 고리
- L
- P
- C
- MS
- C고리 MS고리가 회전을 일으키는 원동력이다.
- 편모의 소단위 단백질 : 플라젤린
- 4개의 고리
- 질주 : 시계 반대방향으로 회전
곤두박질 : 시계 방향으로 회전 - 편모운동
- 고정자(= MotA + MotB) 중심엔 축차(= MS고리 + C고리)가 있다.
- MotA와 MotB가 고정자로 작용한다.
- MotA와 FliG는 톱니처럼 맞물려있다.
- MotA가 돌면 그 힘에 의해서 FliG가 돈다.
- FliG는 뭐냐? C 고리에 있는 단백질이다.
- 그래서 C, MS고리가 돌면서 전체가 돈다.
- 편모 회전력 = 양성자 동력
- 세포질 밖의 많은 수소이온이 들어오면서 회전한다.
- 수소이온 → MotA,MotB 복합체(통로) → MotA,B 복합체 회전 → C고리 회전 → MS고리 회전 → 편모 전체 회전
- 주화성운동 : 농도 기울기에 따른다.
- 질주 : 방향성
- 곤두박질 : 무방향성
- 포자낭이 전포자가 성장할 수 있게 도와준다.
결국 전포자가 최종포자가 된다.
미생물학 3장 (1) - 진핵세포의 구조
- 세포골격 3가지
- 미세소관 ( 20nm) : 튜블린단백질 #방추사 형성
- 중간섬유 ( 10nm) : 케라틴 #세포소기관 제자리에 고정 ex)핵라미나
- 미세섬유 ( 5nm ): 액틴단백질 #원형질 유동
- 골지체는 딕티오솜 구조로 이루어져 있다. ( 납작한 시스테나 층 )
단백질은 소포체에서 먼저 합성되고 골지체에서 포장된다. - 유비퀴틴 : unfolding 된 단백질을 분해하는 프로테아솜에게 신호를 주는 표지자
- 26S 프로테아솜 : unfolding 단백질을 분해한다. 유비퀴틴은 재생산된다.
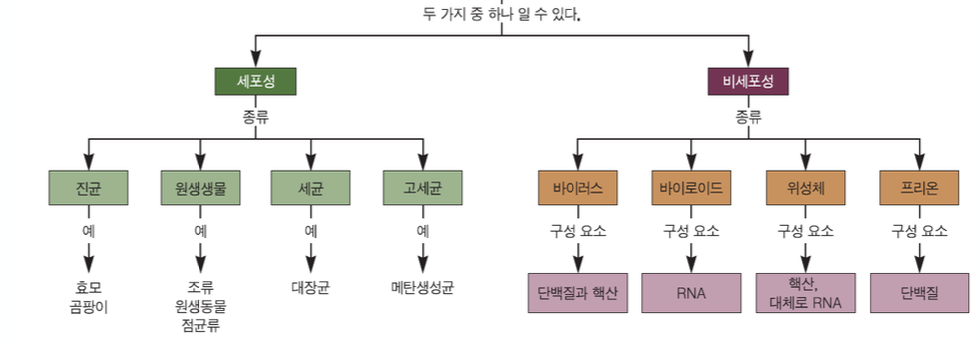
- 우즈 폭스 분류체계 : 3영역 분류
- 세균 : 원핵미생물
- 고세균 : 원핵미생물
- 진핵생물 : 원생생물, 진균, 식물, 동물
미생물학 3장 (2) - 진핵세포의 세포내흡입
- 세포내흡입 : 세포막이 함입되면서 흡입
- 음세포작용(피노솜) : 작은 입자 흡입
- 식세포작용(파고솜) : 큰 입자 흡입 , 파고솜만이 direct하게 라이소좀과 결합한다.
- 클라스린의존성(클라스린피복구) : 호르몬, 콜레스테롤 흡입
- 카베올린의존성(카베오솜) : 신호 전달이 되어야 흡입을 시작한다.
- 카베오솜 → 초기엔도솜과 융합 → 후기엔도솜과 융합 → 라이소좀과 융합
- 자가소화 (autophage) : 미토콘드리아가 파괴될 때 **
- 진핵세포의 리보솜 : 80S Ribosome
- 60S + 40S
- 60S = 5S / 5.8S / 28S rRNA + 50개 단백질
- 40S = 18S rRNA + 30개 단백질
- 세균의 리보솜 : 70S Ribosome
- 50S + 30S
- 50S = 5S / 23S rRNA + 34개 단백질
- 30S = 16S rRNA + 21개 단백질
- 진핵세포의 미토콘드리아의 리보솜 : 70S Ribosome (80S Ribosome 아니다)
- 수소발생체 : 무산소성 원생생물에 있는 세포소기관
산소가 없는 환경에서 해당과정을 거쳐 ATP를 만든다. - 편모와 섬모의 미세구조
- 9+2 구조
- 둘레 미세소관 9쌍 + 중심 미세소관 2개 = 총 20개의 미세소관
- 둘레에 있는 이중미세소관 = 원형 + 반원형 구조
- 기저체의 구조
- 9+0 구조
- triplet 구조 = 3 x 9 = 총 27개의 미세소관
- 세균 / 고세균 / 진핵생물
인트론 X 드물게 O O >> 고세균이 세균과 진핵생물의 중간다리를 한다고 생각됨
펩티도글리칸 O X X
리보솜 70S 70S 80S
세균, 고세균 = 원핵
진핵생물 = 진핵 - 에스테르 결합 = 알코올기 + 카복실기 (축합반응) : 고세균에 없고 진핵생물에 있음
- 에테르 결합 = 알코올기 + 알코올기 (축합반응) : 고세균에만 있음 >> 더 견고
미생물학 4장 (1) - 바이러스의 구조
- ATV : 고세균바이러스 : 고온에서 숙주 밖에 있을 때 꼬리가 관찰된다.
- 비리온 : 완전한 입자형태가 갖추어진 바이러스 입자
- 뉴클레오캡시드 : 핵산 + 단백질 껍질 (막이 없다)
- 캡시드 : 핵산보호
- 핵산 번역 >> 단백질 생성 >> 프로토머 >> 캡소머
- 프로토머가 모여서 캡소머가 된다.
- 스파이크 단백질 : 외피가 있는 바이러스의 막에 있는 막단백질
수용체에 붙기 위해 작용 - 캡시드 모양 2가지
- 정이십면체 캡시드
- 나선형 캡시드 : RNA를 나선형으로 감싸는 캡시드
- 바이러스의 핵산 종류
- DNA
- RNA
- double strand
- single strand
- + : 단백질을 바로 번역할 수 있다.
- - : +로 된 후 단백질을 번역할 수 있다.
- 코로나 바이러스
- 스파이크 단백질이 ACE2 라는 수용체에 가서 붙는다.
- 코로나 바이러스의 분류
- 목 : Nidovirales
- 과 : Coronaviridae
- 속 : Betacoronavirus
- 종 : 2019-nCov
미생물학 4장 (2) - 인플루엔자 바이러스
- 코로나 바이러스
- ACE2 : angiotensin 자르는 효소, 코로나의 수용체 역할 하는 단백질
- 스파이크 단백질
- 당단백질
- sugar chain : 인체의 면역시스템 피해가는 기작
- ACE2 의 binding interface에 붙는다.
- ACE2 와 binding 일어나는 부위 : 스파이크 단백질의 윗부분
- 원형질막과 융합이 일어나는 부위 : 스파이크 단백질의 뿌리부분
- 인플루엔자 바이러스
- 외피 O
- single strand RNA가 분절되어 있다 = 분절유전체
- single strand RNA를 나선형캡시드가 감싸고 있다.
- 나선형캡시드 -> 유연성이 높다 -> 형태다형성
- 스파이크 단백질 2가지
- 헤마글루티닌 (H) : 적혈구 응집소 : 적혈구를 응집시킬 수 있다. (항원-항체)
- 시알산 결합시키는 단백질
- 세포에 들어갈 때 필요하다.
- 뉴라미니데이스 (N)
- 시알산 분해하는 단백질
- 세포에서 방출될 때 필요하다.
- 기질 단백질 (M) : 외피 안쪽에 있다.
- ( H,N ( M ( 나선형캡시드) M ) H,N ) 구조
- 헤마글루티닌 (H) : 적혈구 응집소 : 적혈구를 응집시킬 수 있다. (항원-항체)
- 아데노 바이러스
- 외피 X
- 정이십면체 뉴클레오캡시드
- 캡시드 스파이크 O (스파이크 단백질 X) : binding 할 때
미생물학 4장 (3) - 박테리오페이즈
- 바이러스의 핵산 ★
- 대부분의 세균바이러스는 dsDNA를 갖는다.
- 대부분의 RNA바이러스는 ssRNA를 갖는다.
- 대부분의 식물바이러스는 ssRNA를 갖는다.
- 인플루엔자, 사스, AIDS는 ssRNA를 갖는다.
- 대부분의 RNA바이러스는 분절유전체를 갖는다.
- 숙주 용균에 필요한 단백질
- 홀린 : 원형질막에 구멍을 내게 한다.
- 라이소자임 : 펩티도글리칸을 분해한다.
- 스파이크 단백질 = 페플로머 라고도 한다.
- 바이러스의 증식
- 흡착 → 침투 → 껍질 벗기 → 합성,복제 → 조립 → 방출
- 흡착 : 바이러스의 스파이크가 세포의 수용체와 결합
- 그람 음성 : 지질다당체가 수용체로 작용
- 그람 양성 : 테이코산, 선모, 편모가 수용체로 작용
- 동물바이러스 : 지질뗏목(lipid raft)가 수용체로 작용
- 침투 : 바이러스가 세포막에 의해서 함입되어 내부로 이동, 소낭 막과 외피가 융합되어서 캡시드가 노출된다.
- 원형질막융합 침투 : 외피 보유바이러스일 때
- endocytosis가 아니다
- non-endosome 융합이다.
- 외피와 원형질막이 융합되고 캡시드만 빠져나온다.
- 세포내흡입 침투(endocytosis) : 외피 보유바이러스일 때
- 스파이크 단백질이 세포막과 융합하여 엔도솜 안으로 바이러스가 들어간다.
- 엔도솜 안쪽의 pH가 낮게 유지되면서 엔도솜 막과 외피가 융합되어서 세포질로 빠져나온다.
- 세포내흡입 침투(endocytosis) : 외피 비보유바이러스일 때
- 엔도솜 안쪽의 pH가 낮게 유지되면 캡시드와 엔도솜이 오픈되면서 핵산만 나온다.
- 원형질막융합 침투 : 외피 보유바이러스일 때
- 합성 : RNA, 스파이크 등 합성
미생물학 4장 (4) - 용균성, 용원성
- 용원성 (lysogenic)
- 숙주가 휴지상태에 있을 때, 파지는 용원상태로 염색체 유지
- Prophage : 파지의 DNA가 세균의 염색체 안으로 들어가버린 상태
- 프로파지 상태로 감염된 숙주 : 용원균
- 중복감염 면역성 : 용원균은 동일한 파지에 의해 재차 감염이 일어나지 않는다
- 그람음성 살모넬라균 : 파지감염이 되면 LPS가 수용체 역할을 못하면서 중복감염 면역성이 생긴다.
- 바이러스 감염의 종류
- 급성감염 : 들어가자마자 용균해서 바이러스를 방출시킨다. 비리온을 방출시킨다.
- 잠복감염 : 유전체는 존재하지만 세포가 죽지않음. 비리온을 방출시키지 않는다.
- 만성감염 : 평생 갖고 간다. 비리온 조금씩 방출
- 암세포 유도 : 바이러스가 암유발 유전자를 가지고와서 우리의 세포가 암으로 변형
- 종양억제유전자인 Rb, p53이 돌연변이가 일어나면 암이 발생
미생물학 4장 (5) - 바이로이드, 프리온
- 바이로이드 : 원형의 단일가닥 RNA
- 바이러스와 달리 핵산이 단백질 껍데기로 싸여있지 않다.
- 단백질이 없는 100% RNA로만 되어있는 상태
- 바이로이드 RNA는 단백질을 암호화하는 유전자가 없다. ****
- 어떠한 바이러스의 도움 없이 감염이 되고 복제도 가능하다. ****
- RNA사일런싱으로 질병 유발 : 바이로이드RNA가 숙주의 mRNA서열에 결합하여 자른다. 숙주의 RNA파괴, 질병유발
- 프리온
- 핵산이 없다.
- 단백질 자체가 감염원이다.
- 기작
- 감염원이 들어온다.
- 정상프리온이 비정상프리온으로 바뀐다.
- 비정상 프리온의 응집화가 일어남 : aggregation
- 비정상 프리온의 섬유화가 일어남 = 길어짐 : fibrilization
- 뇌세포 사멸
- 크로이츠펠트-야콥병 (CJD) : 인간 광우병
- Stanly Pruisner - “Protein only hypothesis”
미생물학 5장 (1) - 미생물의 생장
- 세균의 증식방법
- 이분법
- 출아
- 중복분열 : 남세균에서 일어난다. ****
- 대장균의 세포분열
- 세포가 복제될 때 레플리솜(대장균 복제에 관여하는 단백질들이 이룬 구조)이 형성된다.
- 복제 후 분리가 되기 전에 이미 복제원점으로부터 복제가 시작되어 진행된다.
- 즉 복제 & 세포질분열 동시에 일어난다.
- 세포질분열 : FtsZ 단백질이 Z고리를 형성하여 세포질 분열한다.
- FtsZ
- 세포골격 단백질
- Z고리를 만드는 단백질
- FtsZ는 원형질막 바로 아래에 생성된다.
- Z고리는 가운데에 몰려 있다.
- Z고리가 형성되면 세포막이 함입되고 격벽이 합성된다.
- MinCDE 단백질 복합체
- Z고리 형성을 억제한다.
- Z고리가 있는 부분에서는 MinCDE 농도가 낮다.
- MinCDE가 없는 부분에 Z고리가 형성 된다.
- MinCDE는 양 끝 쪽에 몰려 있다.
- 디비솜
- 세포 분할을 담당하는 단백질 복합체
- FtsN 단백질
- 수축 개시를 알린다.
- 세포벽 합성, 펩티도글리칸 합성
- 펩티도글리칸 : NAM-NAG 반복
- 박토프레놀 : 펩티도글리칸 합성을 위해 절대적으로 필요한 지질
- 세포질에서 합성된 것을 밖으로 내보내는 지질 운반체
- 박토프레놀-NAM-NAG 형성
- 자가용해효소(autolysin)
- NAG-NAM 글라이코시드 결합을 절단한다.
- 새로운 펩티도글리칸층을 만들기 위해
- autolysin은 디비솜에 있다.
- MreB : 액틴 유사체
- 세포길이와 지름을 결정
- 막대균이 길쭉하게 유지할 수 있는 이유
- MreB가 없는 간균 -> 구형으로 바뀜
- 구균 : MreB 발현 X
- FtsZ
미생물학 5장 (2) - 환경요인에 대한 미생물의 반응
- 용질과 수분 활성도에 대한 미생물의 반응
- 저장액에 들어가면
- 방어기작이 나타난다.
- MS채널이 열린다.
- 고장액에 들어가면
- 원형질막, 세포벽이 분리되는 원형질분리가 일어난다.
- 방어기작이 나타난다 : 세포내를 고농도로 유지
- 내삼투성 (호염성) : 수분활성도가 0.6에서 최적이 된다. **
- 저장액에 들어가면
- pH에 따른 미생물의 반응
- 호중성 : 주변이 중성
- pH 5.5보다 밑으로 떨어지면 내산반응 일어난다.
- 내산반응 : 산을 견디며 중성을 유지하려고 함
- 세포질의 양성자를 방출하며 자신의 산도를 높인다.
- 그러기 위해서 양성자수송 ATP 가수분해효소 작용 일어난다. **
- pH 4.5보다 밑으로 떨어지면 산충격단백질 합성 유도
- 산충격 단백질 : 단백질에서의 산성을 복구하는 복구단백질
- pH 5.5보다 밑으로 떨어지면 내산반응 일어난다.
- 호산성 : 주변이 산성
- 자기의 pH를 중성으로 유지하려고 한다.
- 세포 내부의 pH가 낮은게 아니다.
- 외부에서 들어오는 수소이온을 계속 밖으로 방출
- 호염기성 : 주변이 염기성
- 자기의 pH를 중성으로 유지하려고 한다.
- 외부의 수소이온을 계속 안으로 끌어들이려고 한다.
- 호중성 : 주변이 중성
- 온도에 따른 미생물의 반응
- 호냉 : 추운 곳에 산다
- 불포화 지방산이 많다 **
- 부동단백질을 이용해 빙점을 낮춘다. **
- 산소농도에 따른 미생물의 반응
- 절대 산소 요구성 : 꼭 산소가 있어야 산다
- 최종 전자 수용체 = O2 **
- + SOD
- + 카탈라아제
- 저농도 산소 요구성 : 꼭 산소가 있어야 산다
- 최종 전자 수용체 = O2 **
- + SOD
- + 카탈라아제
- 조건부 산소 비요구성 : 무산소, 유산소 모두 사용하며 생존
- 내기성 산소 비요구성 : 산소 사용 불가
- 절대 산소 비요구성 : 산소가 없어야 산다
- 포르피로모나스 진지발리스 **
- - SOD
- - 카탈라아제
- SOD와 카탈라아제는 산소의 독성을 막아준다.
- 절대 산소 요구성 : 꼭 산소가 있어야 산다
- 활성산소 (ROS) : 독성산소유도체 **
- 단백질, 지질, 핵산 손상
- 초과산화물 라디칼
- 과산화수소 (H2O2)
- 수산화라디칼 (OH-)
- SOD : 초과산화물 라디칼 제거하는 효소
- 카탈라아제 : 과산화수소를 제거하는 효소
- 단백질, 지질, 핵산 손상
- 생물막 (Biofilm) : 고착미생물 공동체 ** 사진참고
미생물학 5장 (3) - 배지에서의 생장
- 세포간 의사소통
- 해양발광세균 : AHL **
- 오징어의 감광기관에서 AHL을 통해 발광유전자가 발현된다.
- 배지 (culture media)
- 지지배지 (영양배지) : 생장을 위한 배지
- 농화배지
- 선택배지
- ex) MSA배지 : NaCl 농도가 높다
- 포도상구균을 선택적으로 자라게 할 수 있다. **
- 분별배지
- 미생물 생장곡선
- 지체기
- 대수기
- 정체기
- 사멸기
- Viable But Non Culturable (VBNC) **
- 살아있는데 실제로 키우면 안자란다.
- 정체기에서 유전적 변형이 일어나서 생장이 멈춰있다.
- 조건이 회복되면 생장이 다시 진행된다.
- 대수기에서의 미생물의 생장 **
- n : 분열 횟수
- t : 배양 시간
- g : 세대 시간 = 한번 분열하는데 걸리는 시간 = t/n
- k : 생장률 상수 = t 시간 동안 분열 횟수 = n/t
- Nt = No x 2^n
미생물학 6장 (1) - 미생물 제어 방법
- 미생물 제어 방법
- 물리적 제재
- 화학 제재
- 기계적 제거 방법
- 생물학적 제재
- 화학적 제재
- 멸균 > 소독 > 방부 **
- 방부 : 생체조직의 미생물을 죽인다.
- 조직에 있는 병원체 억제, 파괴
- 소독 : 생체조직이 아닌 미생물을 다룬다.
- 감염원 크기 축소 조치
- 멸균 : 자를 수 있는 균을 0으로 만든다.
- 살아있는 감염원 완전 파괴
- 미생물의 사멸 양상 D값, Z값
- D = 미생물 90%가 죽을때까지의 시간 **
- D가 짧으면 짧을수록 멸균 장치가 강하다.
- Z = D값이 10에서 1이 되는 온도차이 **
- Z가 작을수록 멸균력이 좋다.
- D = 미생물 90%가 죽을때까지의 시간 **
- 기계적 제거 방법
- HEPA filter : 고효율입자 공기 필터
- 촘촘해서 바이러스까지 제거 가능
- 물리적 제재
- 습열 멸균법
- 고압습윤멸균기 : 121℃, 15 파운드 압력 **
- 틴들법 : 내생포자 제거 **
- 방사선 처리법 : DNA 복제와 전사에 손상
- 비열멸균법
- 내생포자는 파괴되는데 바이러스는 예외이다. **
- 건열 멸균 : 멸균속도 느리다 **
- 습열 멸균 : 멸균속도 빠르다 ** 습해서 움직이면 애들 잘보임
- 습열 멸균법
- 살균제의 활성
- 페놀계 화합물 : 크레솔, 자이레놀 (세포막파괴) **
- 알코올 : 세균, 진균에 효과, 포자에는 무효과 **
- 요오드, 염소 : 단백질 산화작용이 강하다. **
- 글루타르알데하이드 : 단백질에 달라붙어서 단백질이 denaturation된다. **
미생물학 8장 - 물질대사
- 자유에너지 변화 = 엔탈피 변화 - T엔트로피 변화
= 총열량 변화 - 무질서도 변화 - 자유에너지
- 계 안에서 반응이 일어날 때 반응을 일으키는데 필요한 에너지
- 엔탈피 변화 : 어떤 반응이 일어날 때 열량이 총체적으로 변한 에너지
- T엔트로피 변화 : 무질서도의 변화
- △Gº' = 표준 자유에너지변화
- △Gº' = -2.303 x R x T x logKeq
- △Gº'가 (-) 이면 Keq는 1보다 크다.
- 에너지 방출이 진행되고 자발적으로 이루어진다.
- △Gº'가 (+) 이면 Keq는 1보다 작다.
- 에너지 흡수가 진행되고 비자발적으로 이루어진다.
- Eo’ : 표준환원 전위
- Eo'(-) 의 의미 : 훨씬 더 전자를 잘 준다. 전자공여체이다.
- Eo'(+) 의 의미 : 훨씬 더 전자를 잘 받는다. 전자수용체이다.
- Gº' = -n x F x △Eo'
- n : 전달된 전자수
- F : 페러데이 상수
- △Eo' = Eo'(수용체) - Eo'(공여체)
- 표준 자유에너지 예제
- NADH에서 O2로 전자 2개가 이동할 경우 표준 자유에너지 변화는?
- -(전달된 전자수) x (페러데이 상수) x (수용체 O2의 Eo' - 공여체 NADH의 Eo')
- -2 x 23.052 x (0.82-(-0.32))
- 미토콘드리아의 NADH 탈수소효소 **
NADH로부터 전자를 빼서 넘겨주는 효소 - NAD+ 에 전자 2개와 양성자 2개가 들어가면
NAD+ = NADH + H- : 양성자는 하나만 받아서 하나가 남는다. - 조효소Q = 유비퀴논 = CoenzymeQ : 전자를 받는다. **
- 헴(heme) : 전자 1개만 운반 **
- 효소반응
- 효소기질 복합체 형성
- 효소가 활성화 에너지를 낮춘다. **
- 전이상태 복합체 형성
- 반응속도 증가
미생물학 9장 (1) - 에너지 공급
- 미생물 분류 기준
- 에너지원 : 빛 사용 / 유기,무기화합물 사용
- 전자원 : 전자주느냐 / 전자받느냐
- 탄소원 : CO2 이용 / 유기탄소 이용
- 산소호흡의 연료공급
- PMF (양성자 구동력)
- SLP (기질수준인산화)
- EMP (Emden-Meyerhof Pathway)
- 해당과정이라고도 부른다.
- 최종산물 : 2 피루브산, 2 ATP, 2 NADH
- 고에너지 물질 2가지 **
- 1,3 - 이인산글리세르산
- 포스포에놀피루브산
- EDP (Entner-Doudoroff Pathway)
- 진핵미생물에서 작동 안하고 그람양성, 그람음성균에서 작동한다.
- 포도당 6-인산 -> 6-포스포글루콘산 **
- 이때 NADP+ 가 필요하다 **
- 최종산물 : 1 ATP, 1 NADH, 1 NADPH (환원력)
- PPP (Pentose Phosphate Pathway)
- 트랜스케톨라아제 ****
- 탄소 2개를 절단해서 붙여주는 효소이다.
- 트랜스알돌레이스 ****
- 탄소 3개를 절단해서 글리세르알데하이드에 붙여주는 효소이다.
- PPP는 NADPH가 많이 만들어진다.
- 그러면서 동화반응의 환원력을 생산하는 주된 경로이다. ****
- 전구대사물질의 생성경로이다. ****
- 전구대사물질 :
- 에리스로스-4-P : 방향족아미노산을 생합성하는 전구물질 ****
- 리보오스-5-P : 핵산을 생합성하는 전구물질 ****
- 트랜스케톨라아제 ****

미생물학 9장 (2) - TCA 회로, 전자전달계
- TCA 회로
- TCA 회로가 일어나는 위치
- 진핵미생물 : 미토콘드리아 기질
- 세균, 고세군 : 세포질 ★
- 과정
- 아세틸CoA (C2) 가 처음으로 TCA회로에 들어간다.
- 아 - 시 - 아 - 숙 - 푸 - 마 - 옥
- 숙시닐-CoA 가 기질수준 인산화로 인해 ATP가 생기면서 숙신산이 된다.
- TCA 회로의 고에너지 화합물 = 숙시닐-CoA : 기질수준 인산화가 일어났기 때문에 ★
- MDP에서의 고에너지 화합물 = 1,3 이인산 글리세르산, 포스포에놀 피루브산 ★
- TCA 회로가 일어나는 위치
- 전자전달계
- 전자전달계 위치
- 진핵미생물 : 미토콘드리아 내막
- 세균, 고세균 : 원형질 막
- 전자전달계 특징
- FMN, FAD, CoQ는 전자와 양성자를 동시에 수송한다.
- FeS는 전자만 수송한다.
- NADH 가 산화되면 10개의 H+ 가 막간공간으로 이동한다. ★
- ATP합성효소는 막간공간에서 4개 H+를 흡입하여 1개의 ATP를 만든다. ★
- 3개의 H+는 ATP합성효소가 돌게 만들고 1개의 H+는 ADP가 활성 site에 붙는데 쓰인다.
- 1개의 NADH는 2.5개의 ATP를 만든다. ★
- FADH2 가 산화되면 6개의 H+가 막간공간으로 이동한다.
- 1개의 FADH2는 1.5개의 ATP를 만든다. ★
- 전자전달계 위치
- 대장균의 전자전달계
- FAD → FeS 과정에서 FeS는 전자만 수송하므로 남은 양성자를 바로 Q에 붙여준다. ★
- Q → QH2 : 전자 2개와 양성자 2개를 받아들임 ★
- 산소공급이 불충분할 때 : bd 가지 ★
- 산소공급이 충반할 때 : bo 가지 ★
- 대장균의 ETC는 총 5가지가 작동한다. ★
- Q회로 ★★
- Coenzyme Q는 여러가지 complex 사이에서 중간자 역할을 한다.
- 양성자 구동력을 만드는 역할
- 전자 하나를 받음 : Q → [1e] → Q-
- 남은 전자 하나와 양성자 2개를 받음 : Q- → [1e, 2개의 양성자] → QH2
- Q회로 : Q → [1e] → Q- → [1e, 2개의 양성자] → QH2 ★
미생물학 9장 (3) - 무산소호흡, 발효, 베타산화
- 합성되는 ATP 계산
- 1 NADH → 2.5 ATP
- 1 FADH2 → 1.5 ATP
- 1 아세틸CoA → 10 ATP
- 포도당 1분자당 해당과정으로 만드는 ATP 양
- 2 NADH(산화적) + 2 ATP(기질수준) → 7 ATP
- 피루브산 1분자당 만드는 ATP 양
- 1 NADH + 3 NADH + 1 FADH2 + 1 ATP(기질수준) → 12.5 ATP
- 아세틸CoA 1분자당 만드는 ATP 양
- 3 NADH + 1 FADH2 + 1 ATP → 10 ATP
- [세포질] 포도당 → 피루브산 : 7 ATP 생성
- [미토콘드리아 내막] TCA 회로 : 25 ATP 생성
- 총 32 ATP 생성
- 무산소 호흡 : 산소가 아닌 다른 물질이 전자를 받는다.
- 탈질작용 : 질소기체를 방출한다. ★
- 최종 전자수용체 : NO3- ★
- NO3- 는 최종적으로 N2 가 된다.
- 단당류 4종류 젖당발효 시
- 포도당
- 만노오스 → 포도당-1-P ★
- 과당
- 갈락토오스
- β 산화 싸이클 : 지질의 이화작용 ★
- 지방산 → β 산화 싸이클 → 탄소 2개씩 방출 ★
- fatty acid가 어떻게 이화 되는가? : β 산화 싸이클
- 아세틸CoA가 만들어지므로 에너지가 어마어마하다.
C16 fatty acid의 16개의 탄소가 β싸이클을 돌고 난 후
8 아세틸-CoA + 7 FADH2 + 7 NADH 가 생기므로
8 아세틸-CoA = 8 x 10 ATP = 80 ATP
7 FADH2 = 7 x 1.5 ATP = 10.5 ATP
7 NADH = 7 x 2.5 ATP = 17.5 ATP
총 108 ATP가 생성됩니다.
106 ATP
미생물학 9장 (4) - 무기영양생물의 연료공급, 광합성
- 탈아미노기 반응 ★
- 무기영양생물의 연료공급
- 연료를 공급하려면 환원력(NADPH)와 ATP가 필요하다. ★
- 무기분자로부터의 전자전달계의 작동
- 무기분자 산화의 3가지 유형
- 수소효소작용 : 수소 가져다가 양성자와 전자로 나눠 : H2 -> 2H+ , 2e-
- 질화작용 : 질소의 산화 : NH4+ → NO2- → NO3-
- 황산화작용 : S → SO4-
- APS는 ATP 합성에 이용된다 : APS + Pi = ADP ★
- 역전자흐름 ★
- 양성자가 주변 세포질에서 세포질로 이동하는 작용
- 전자전달 정방향의 역방향으로 작동된다.
- 이는 NADH 를 생성하기 위함이다.
- 광합성
- 그라나를 싸고 있는 막 구조 : 틸라코이드
- 나머지 액체로 채워진 부분 : 스트로마
- 빛에 직접 contact 하는 부분 : 틸라코이드 막
- 4개의 H+ 소비 = 3개 H+ (ATP합성효소 120도 회전) + 1개의 H+ (ADP + Pi 결합)
- 광인산화 과정
- 빛이 P700 때림
- P700의 전자가 붕 뜬다.
- 전자가 붕 떴으므로 빈자리를 채워줘야 한다.
- P680의 전자가 붕 뜬다.
- 전자가 붕 떴으므로 빈자리를 채워줘야 한다.
- 물이 분해되면서 H가 가지고 있던 전자가 빈자리를 채워준다.
- 물의 분해는 OEC(산소방출복합체)가 해준다
- Oxgen Evolving Complex
- 물이 분해되면 산소는 방출되지만 수소는 방출되지 않는다.
- 수소를 H+와 전자로 나누는 역할을 한다.
- H+ 는 틸리코이드 안에 쌓이게 된다.
- PMF는 틸라코이드 내강에 생긴다.
- H+는 ATP 합성효소를 통해 빠져나가면서 ATP가 만들어진다.
- P700의 붕 뜬 전자는 NADP+가 받아서 NADPH가 만들어진다.
- P700
- 순환적 광인산화 : NADPH 생성 X, ATP 생성 O
- 비순환적 광인산화 : NADPH 생성 O, ATP 생성 O
- P680
- 순환적 광인산화 할 수 없다.
- 페러독신 : 비순환, 순환 경로를 결정하는 인자.
- 비순환적 광인산화
- 물 2분자 분해 → 산소 1분자 생성 → 전자 4개 방출 → H+ 4개가 틸라코이드 안에 생성
- 전자 4개 운반 → 스트로마에서 H+ 4개 공급 → 전자 운반시 이용되고 틸라코이드 안에 H+ 4개 생성
- 총 8개의 H+ 가 PMF 발휘 → 2 ATP 생성
- 결과 : 2 NADPH + 2 ATP
- 녹색세균의 산소 비발생 광합성
- 클로로솜 : 세균엽록소를 가지고 순환적 인산화를 한다.
- 광계 1만 존재한다.
- 로돕신 광영양 ★
- 고염분일 때 로돕신의 구조가 변하면서 양성자를 방출하며 주변세포질에 PMF가 생기면서 ATP를 생산한다.
미생물학 11장 (1) - 전사
- 폐렴균 형질전환 실험
- 그리피스 (1928)
- 사멸된 S형 균주 + 살아있는 R형 균주 = 살아있는 S형 균주 + 살아있는 R형 균주
- DNA가 유전물질일 것이다.
- 에이버리
- RNA와 단백질을 파괴해도 형질전환이 일어났다.
- DNA를 파괴하니 형질전환이 일어나지 않았다.
- 형질전환을 일으키는 요소는 DNA이다.
- 허스 & 체이스 (1952)
- 박테리오파지를 35S로 표지된 껍질 단백질, 32P DNA를 표지하고 세균에 감염
- 세균 안으로 들어간 것은 단백질이 아니라 DNA이다.
- 유전물질은 단백질이 아니다 를 증명
- 왓슨 & 크릭 (1953)
- DNA 이중나선 발견
- 그리피스 (1928)
- E coli
- 세균 염색체는 환형이다. ★
- oriC 라는 복제원점에서부터 복제가 시작된다.
- 세타구조 : 전체 염색체 중 30%가 복제된 상태
- DNA 중합효소 lll = 레플리솜 (replisome) ★
- 핵심효소
- 베타 클램프 : DNA를 핵심효소에 붙게 해준다.
- 클램프로더 : 베타 클램프를 DNA에 올려 놓아준다.
- 타우 : 핵심효소를 잡는다.
- Dna A : 복제원점인 oriC에 가장 먼저 붙는다. 헬리케이스를 대리고 온다.
- Dna B : 헬리케이스 그 자체
- Dna C : 헬리케이스를 DNA에 붙여준다.
- 헬리케이스 : 이중가닥을 분리한다. (ATP 가수분해)
- SSB : 이중가닥이 단일가닥이 되면 그 단일가닥을 안정화 시킨다. 풀린 상태 유지
- topoisomerase ll : 이중가닥의 안풀린 부분(긴장 = supercoil)을 풀어준다. 대장균에서는 gryase라고 불린다.
- pirmase : RNA 프라이머를 합성한다.
- primosome : primase + 보조단백질
- 오카자키 조각 : 연속으로 합성되지 않고 불연속으로 합성된다.
- 선도가닥 : 연속적으로 합성되는 가닥
- 지연가닥 : 불연속적으로 합성되는 가닥
- DNA 중합효소 l : 지연가닥의 RNA 프라이머를 제거한다. ★
- 지연가닥 선도가닥 합성 과정 ★
- 클램프로더가 베타클램프를 잡아서 RNA 프라이머가 있는 곳에 안착시킨다.
- 베타클램프가 DNA를 감싸면서 합성이 진행된다.
- 합성이 끝나면 타우가 핵심효소를 잡아서 분리한다.
- DNA 중합효소 l 이 오카자키 조각의 프라이머를 제거하고 그 제거된 부분을 DNA 합성을 통해 채운다.
- 오카자키 절편 간의 간극은 ligase가 채운다.
- DNA 중합효소 lll 의 교정 : 잘못된 염기서열이 합성되었을 때 (G가 들어가야 되는데 A가 들어감)
- 3' -> 5' 핵산말단분해 활성이 일어난다. ★
- 말단의 1개의 염기를 제거한다.
- 복제 종결
- 연쇄형 염색체 : 위상이성질화효소 작용
- 이량체 염색체 : XerCD 재조합효소 작용 ★
- 전사 : DNA로부터 RNA를 만드는 과정
- 암호가닥 : 5' -> 3'
- 주형가닥 : 3' -> 5' : 전사가 되는 가닥
- 프로모터 (+35 ~ -10)
- -10 지점 : RNA 중합효소 결합부위 : 프립나우 상자
- TATA box가 있다. 수소결합이 약한 부위
- 이 부위가 열려야 전사가 시작된다.
- -10 지점 : RNA 중합효소 결합부위 : 프립나우 상자
- 선도서열
- 선도서열의 +1 지점 = 전사가 시작되는 지점 ★
- 샤인 달가노 서열 을 가지고 있다.
- 번역이 시작되는 서열
- 16s rRNA 의 상보서열을 가지고 있다.
- 16s rRNA는 30S에 존재한다. ★
- 선도서열의 끝나는 부위에 AUG가 붙어서 전사가 개시된다.
- 암호화부위 : 단백질로 번역이 되는 부위, 흔히 말하는 유전자
- 후방서열
- 종결자
- 프로모터 (+35 ~ -10)
- 다중 시스트론 mRNA : 개시코돈 종결코돈이 두개 이상 있는 mRNA (세균, 고세균) ★
단일 스스토론 mRNA : 개시코돈 종결코돈이 하나 (진핵 미생물)
미생물학 11장 (2) - 전사종결, 번역
- -10 서열 = Pribnow box = TATA box
- A와 T가 많아서 쉽게 열린다.
- 열리면서 전사가 시작된다.
- 전사 종결
- 인자비의존적 종결 ★
- 단백질의 필요가 없이 DNA 서열로 종결이 가능하다
- 역반복 서열 : A rich 서열 ★
- A가 많으면 수소결합이 약하다.
- 줄기와 고리구조가 생기면서 끌어당기는 힘으로 작용하여 떨어져나가기 쉬운 모양이 된다.
- RNA 중합효소의 활성이 억제된다.
- 인자의존적 종결 ★
- 로인자가 rut sequence에 붙는다.
- 로인자는 헬리케이스 활성을 통해 RNA,DNA 혼성체가 분리된다.
- 인자비의존적 종결 ★
- 번역 (Translation)
- 암호화 부위에 있는 염기는 3개씩 코돈을 이룬다.
- tRNA의 안티코돈이 코돈을 읽는다.
- tRNA에 붙어있는 아미노산을 연결시키기 위함이다.
- 아미노산을 지정하는 코돈 : 61가지 = 64 - 3(종결코돈)
- 암호의 중복성 : 동일한 아미노산의 코돈이 여러개인 것
- 동요현상 (wobble) : 느슨하다 ★
- 코돈의 3번째 염기는 안티코돈과 염기쌍을 형성할 때 느슨하게 연결된다.
- 코돈의 U와 안티코돈의 G가 염기쌍을 형성할 수 있게 된다.
- 실제로는 45개의 tRNA가 존재하는데, 어떤 tRNA는 하나 이상의 코돈과 결합해야함을 의미하기 때문에 이를 동요현상으로 설명할 수 있다.
- ex) 4개의 글리신 코돈을 번역하는 데에는 3개의 tRNA 안티코돈만 필요하다.
- Polysome : 전사와 번역이 동시에 되는 것
- 아미노아실-tRNA 합성효소 : 아미노산과 tRNA를 붙여준다.
- fMet-tRNA : 개시 tRNA : 메티오닌이 붙어있다.
- rRNA의 3가지 기능
- 리보솜의 전체 구조를 형성한다.
- 30S 소단위의 16s rRNA : 16s rRNA에는 샤인-달가노서열(번역 할 때)이 붙는 자리이다. ★
- 50S 소단위의 23s rRNA : 23s rRNA에는 펩타이드 결합을 촉매하는 라이보자임 역할을 하는 부분이다. ★
- 번역과정 ★
- IF-2 : 30S 소단위의 P자리에 fMet-tRNA를 장착한다.
- EF-Tu : A자리에 아미노아실-tRNA를 장착한다.
- 50S 소단위의 23 rRNA(라이보자임) : P자리와 A자리의 아미노산을 연결한다.
- 21번째 아미노산 : 셀레노시스테인
- 22번째 아미노산 : 피로라이신
- Chaperone : 단백질 folding을 도와준다
- GrpE : 샤페론 cycle을 돌리기 위해, ADP를 떨어뜨리고 ATP를 장착시켜주는 역할 ★
미생물학 12장 (1) - 젖당오페론, 트립토판오페론, Riboswitch
- 젖당오페론
- 조 - 프 - 작 - 구
- 오페론 = 프로모터 + 작동유전자 + 구조유전자 (조절유전자 포함 X)
- 조절유전자 = 억제자를 암호화하는 유전자
- 트립토판오페론
- 전사 활성
- trpR 억제자는 트립토판이 없으면 작동유전자에 붙을 수 없다.
- 전사 억제
- trpR 억제자에 트립토판이 붙으면 작동유전자에 붙을 수 있다.
- trpL(전사약화서열) : 전사가 시작되어도 멈출 수 있다.
- 트립토판이 많을 때
- 3번-4번 고리가 연결되면서 전사종결고리가 만들어진다.
- 트립토판 생합성 유전자의 전사가 멈춘다.
- 트립토판이 없을 때
- 3번-4번 고리가 연결되지 못하고, 2번-3번 고리가 연결되면서 전사종결고리가 만들어지지 않는다.
- 트립토판 생합성 유전자의 전사가 이루어진다.
- 트립토판이 많을 때
- 전사 활성
- Riboswitch ★★
- mRNA 선도서열 내 특정서열의 folding
- ribO : 리보플라빈 오페론
- 리보플라빈이 대사가 되면 FMN이 된다.
- FMN은 rfn 박스에 가서 달라붙는다.
- 그러면 3차구조가 달라진다.
- 고리가 없어지면서 UUUU region 앞에 새로운 고리가 형성된다.
- 정리
- mRNA 전사가 끝까지 지속되면 ribO가 작동하여 리보플라빈이 합성된다.
- 리보플라빈이 대사물질로 변하여 rfn박스에 붙는다.
- 그러면 3차 구조를 변형시킨다.
- 종결자 고리가 만들어지면 리보플라빈을 만들지 않는다.
- 이중 영양 생장 : 포도당을 먼저 쓰고, 젖당을 쓴다.
- 이화물 오페론
- CAP를 활성화 시키려면 cAMP가 필요하다.
- 포도당이 많다 >> ATP 많다, cAMP 적다 >> CAP 비활성화 >> 젖당오페론 비활성화
- 포도당이 적다 >> ATP 소모, cAMP 많다 >> CAP 활성화 >> 젖당오페론 활성화
- 젖당 오페론 기작
- (포도당 X, 젖당 X)
- 조절유전자에서 억제자가 만들어진다.
- 그 억제자가 작동유전자에 붙는다.
- 포도당이 없으면 ATP도 없어지고, cAMP가 많아진다
- cAMP는 CAP 단백질을 활성화시킨다.
- 그러나 억제자가 전사를 불활성화 시켜서 젖당 분해효소가 만들어지지 않는다.
- (포도당 O, 젖당 X) : 가장 젖당 분해 효소가 안 만들어질 때
- 조절유전자에서 억제자가 만들어진다.
- 그 억제자가 작동유전자에 붙으면서 전사가 불활성된다.
- 젖당 분해 효소가 만들어지지 않는다.
- (포도당 X, 젖당 O) : 가장 젖당 분해 효소가 잘 만들어질 때
- 조절유전자에서 나온 억제자가 만들어진다.
- 그 억제자에 젖당이 붙으면서 작동유전자에 젖당+억제자가 붙지 않는다.
- RNA중합효소가 프로모터에 붙는 것을 CAP가 도와준다.
- 포도당이 없다면 ATP -> ADP -> cAMP 과정이 일어난다.
- cAMP는 CAP 단백질을 활성화 시킨다.
- RNA중합효소가 프로모터에 붙으면서 전사가 활성화 된다.
- 젖당 분해 효소가 만들어진다.
- (포도당 O, 젖당 O)
- 조절유전자에서 나온 억제자가 만들어진다.
- 그 억제자에 젖당이 붙으면서 작동유전자에 젖당+억제자가 붙지 않는다.
- 포도당이 있다면 cAMP -> ADP -> ATP 이 과정이 일어난다.
- cAMP의 농도가 낮아지면서 CAP가 활성화되지 않는다.
- RNA 중합효소가 프로모터에 붙지 않는다.
- 젖당 분해 효소가 만들어지지 않는다.
- 포도당을 다 사용하면 3번 메커니즘에 따라서 젖당 분해 효소가 만들어진다.
'🧬 Bio > 미생물학' 카테고리의 다른 글
| 미생물학 12장 (2) (6) | 2020.06.20 |
|---|---|
| 미생물학 12장 (1) - 젖당오페론, 트립토판오페론, Riboswitch (0) | 2020.06.17 |
| 미생물학 11장 (2) - 전사 종결, 번역 (0) | 2020.06.12 |
| 미생물학 11장 (1) - 전사 (0) | 2020.06.11 |
| 미생물학 9장 (4) - 무기영양생물의 연료공급, 광합성 (0) | 2020.06.03 |