
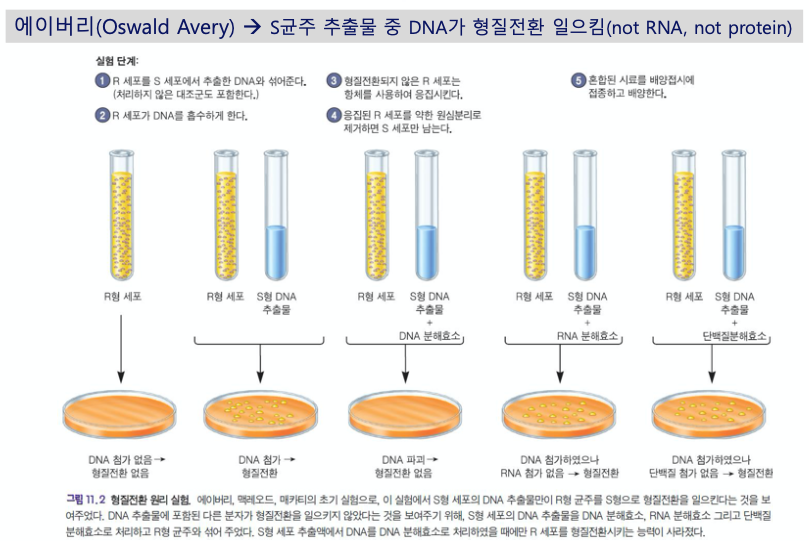
- 폐렴균 형질전환 실험
- 그리피스 (1928)
- 사멸된 S형 균주 + 살아있는 R형 균주 = 살아있는 S형 균주 + 살아있는 R형 균주
- DNA가 유전물질일 것이다.
- 에이버리
- RNA와 단백질을 파괴해도 형질전환이 일어났다.
- DNA를 파괴하니 형질전환이 일어나지 않았다.
- 형질전환을 일으키는 요소는 DNA이다.
- 허스 & 체이스 (1952)
- 박테리오파지를 35S로 표지된 껍질 단백질, 32P DNA를 표지하고 세균에 감염
- 세균 안으로 들어간 것은 단백질이 아니라 DNA이다.
- 유전물질은 단백질이 아니다 를 증명
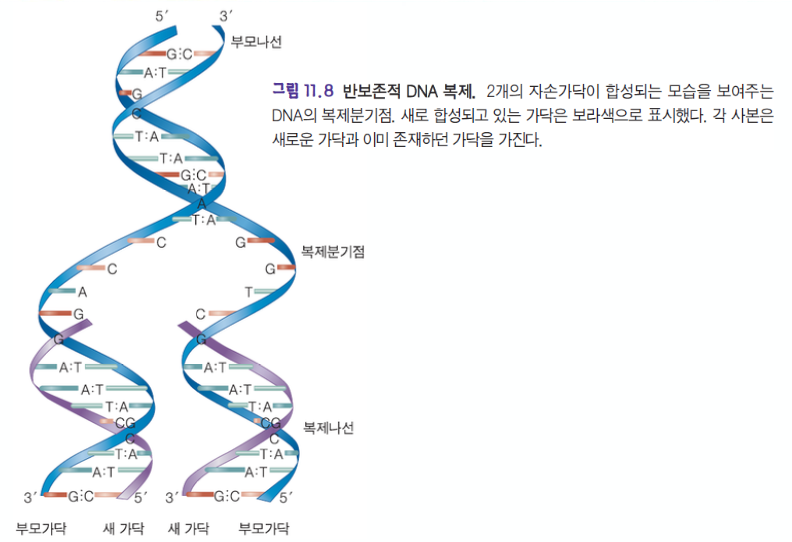
- 왓슨 & 크릭 (1953)
- DNA 이중나선 발견
- 그리피스 (1928)
- E coli
- 세균 염색체는 환형이다. ★
- oriC 라는 복제원점에서부터 복제가 시작된다.
- 세타구조 : 전체 염색체 중 30%가 복제된 상태
- DNA 중합효소 lll = 레플리솜 (replisome) ★
- 핵심효소
- 베타 클램프 : DNA를 핵심효소에 붙게 해준다.
- 클램프로더 : 베타 클램프를 DNA에 올려 놓아준다.
- 타우 : 핵심효소를 잡는다.
- Dna A : 복제원점인 oriC에 가장 먼저 붙는다. 헬리케이스를 대리고 온다.
- Dna B : 헬리케이스 그 자체
- Dna C : 헬리케이스를 DNA에 붙여준다.
- 헬리케이스 : 이중가닥을 분리한다. (ATP 가수분해)
- SSB : 이중가닥이 단일가닥이 되면 그 단일가닥을 안정화 시킨다. 풀린 상태 유지
- topoisomerase ll : 이중가닥의 안풀린 부분(긴장 = supercoil)을 풀어준다. 대장균에서는 gryase라고 불린다.
- pirmase : RNA 프라이머를 합성한다.
- primosome : primase + 보조단백질
- 오카자키 조각 : 연속으로 합성되지 않고 불연속으로 합성된다.
- 선도가닥 : 연속적으로 합성되는 가닥
- 지연가닥 : 불연속적으로 합성되는 가닥
- DNA 중합효소 l : 지연가닥의 RNA 프라이머를 제거한다. ★
- 지연가닥 선도가닥 합성 과정 ★
- 클램프로더가 베타클램프를 잡아서 RNA 프라이머가 있는 곳에 안착시킨다.
- 베타클램프가 DNA를 감싸면서 합성이 진행된다.
- 합성이 끝나면 타우가 핵심효소를 잡아서 분리한다.
- DNA 중합효소 l 이 오카자키 조각의 프라이머를 제거하고 그 제거된 부분을 DNA 합성을 통해 채운다.
- 오카자키 절편 간의 간극은 ligase가 채운다.
- DNA 중합효소 lll 의 교정 : 잘못된 염기서열이 합성되었을 때 (G가 들어가야 되는데 A가 들어감)
- 3' -> 5' 핵산말단분해 활성이 일어난다. ★
- 말단의 1개의 염기를 제거한다.
- 복제 종결
- 연쇄형 염색체 : 위상이성질화효소 작용
- 이량체 염색체 : XerCD 재조합효소 작용 ★
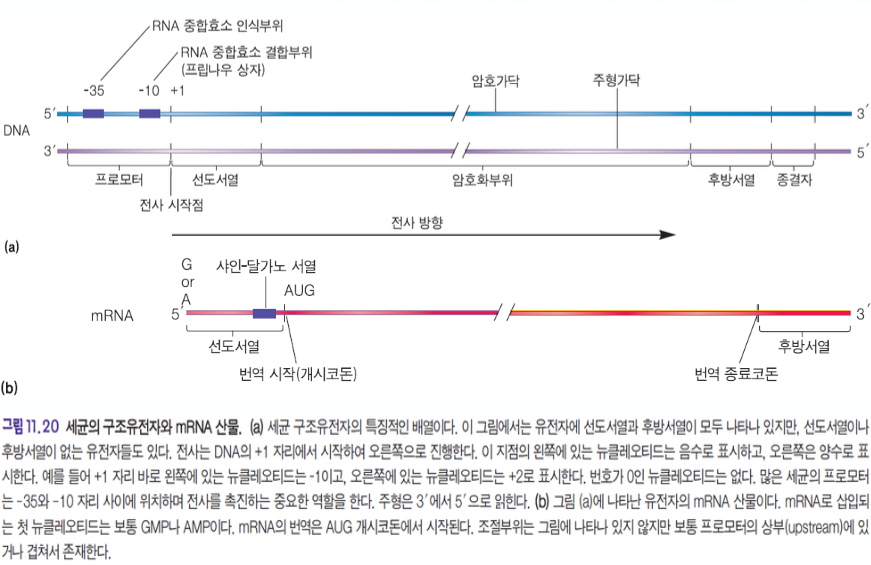
- 전사 : DNA로부터 RNA를 만드는 과정
- 암호가닥 : 5' -> 3'
- 주형가닥 : 3' -> 5' : 전사가 되는 가닥
- 프로모터 (+35 ~ -10)
- -10 지점 : RNA 중합효소 결합부위 : 프립나우 상자
- TATA box가 있다. 수소결합이 약한 부위
- 이 부위가 열려야 전사가 시작된다.
- -10 지점 : RNA 중합효소 결합부위 : 프립나우 상자
- 선도서열
- 선도서열의 +1 지점 = 전사가 시작되는 지점 ★
- 샤인 달가노 서열 을 가지고 있다.
- 번역이 시작되는 서열
- 16s rRNA 의 상보서열을 가지고 있다.
- 16s rRNA는 30S에 존재한다. ★
- 선도서열의 끝나는 부위에 AUG가 붙어서 전사가 개시된다.
- 암호화부위 : 단백질로 번역이 되는 부위, 흔히 말하는 유전자
- 후방서열
- 종결자
- 프로모터 (+35 ~ -10)
- 다중 시스트론 mRNA : 개시코돈 종결코돈이 두개 이상 있는 mRNA (세균, 고세균) ★
단일 스스토론 mRNA : 개시코돈 종결코돈이 하나 (진핵 미생물)
폐렴균 형질전환 실험 : 그리피스 (1928)

S형 균주 : 병원성
R형 균주 : 비병원성
사멸된 S형 균주 + 살아있는 R형 균주 = 살이있는 S형 균주 + 살아있는 R형 균주
에이버리
DNA를 파괴하니까 형질전환이 일어나지 않았다.
RNA와 단백질을 분해해도 형질전환이 일어났다.
따라서 형질전환을 일으키는 요소는 DNA 이다.
허시 & 체이스 (1952)

안으로 들어간 것은 단백질이 아니라 DNA이다.
유전물질이 단백질이 아니다라는 증거를 제시했다.
왓슨 & 크릭 (1953)

DNA 이중나선 발견

Ecoli
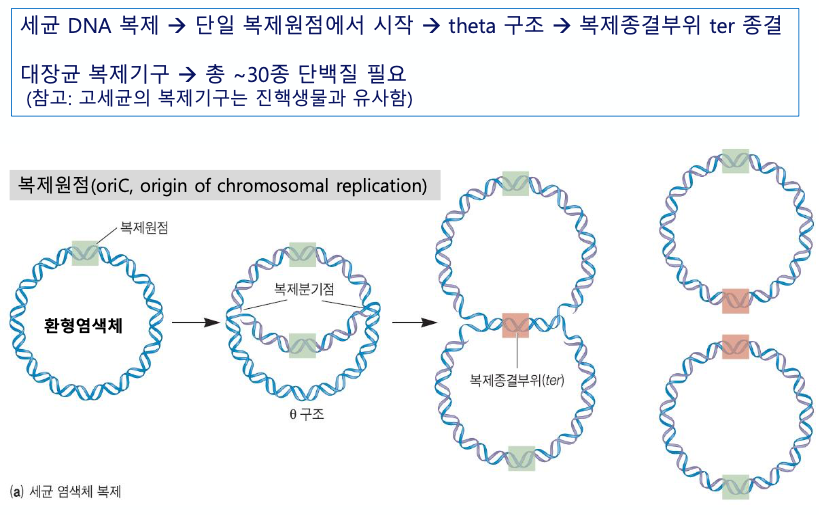
세균의 염색체는 환형이다. ★★★★★★★
복제원점에서부터 복제가 시작된다.
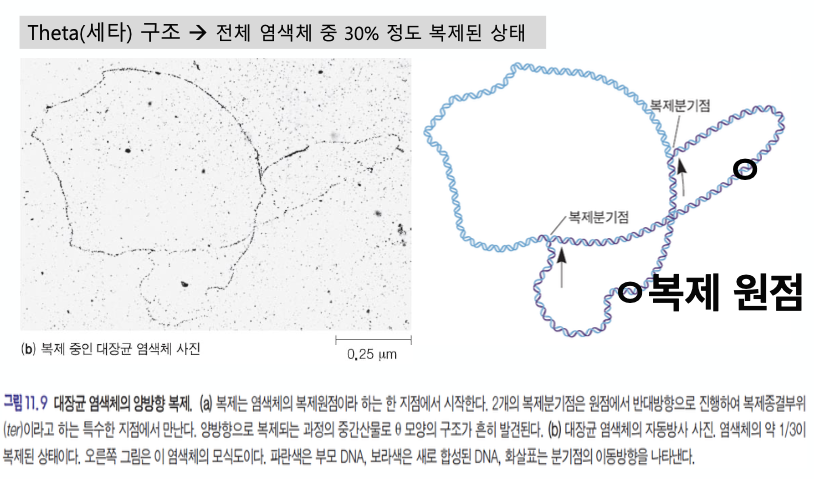
복제 원점은 하나였는데 떨어져서 복제되어 가고 있는 중간단계의 물질이다.
DNA 중합효소 lll
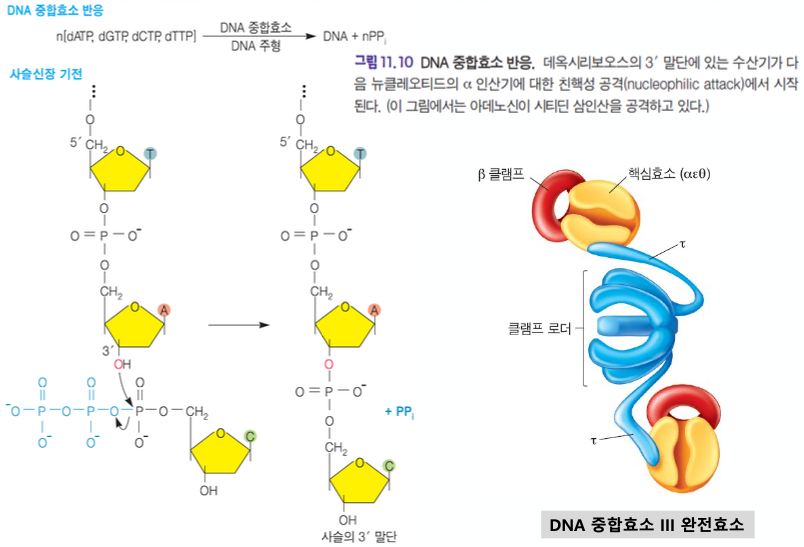
복제하는 효소는 DNA중합효소3 이다.

핵심효소
베타 클램프 : DNA를 핵심효소에 붙게 해준다.
클램프로더 : DNA에 베타클램프를 올려 놓아준다.
타우 이량체 : 2개의 핵심효소가 클램프로더에 잡게 해준다.

DNA 중합효소 lll = 레플리솜 ★★★★★★★
Dna A : 복제원점인 oriC에 제일 먼저 붙는다. 얘가 붙으면서 헬리케이스를 대리고 온다.
Dna B : 헬리케이스
Dna C : 헬리케이스를 DNA에 붙여준다.
헬리케이스 : ATP 가수분해 하면서 이중나선의 가닥을 분리한다.
SSB : 이중나선이 분리되면 단일가닥이 생기는데 그 단일가닥을 안정화시킨다.
toposiomerase ll : 이중나선의 DNA를 풀게되면 아직 안풀린 부분(긴장 = supercoil)이 생긴다. 그것을 풀어준다.
대장균에서는 gyrase라고 불리운다.
primase : RNA 프라이머를 합성한다.
primosome : primase + 보조단백질
Dna B인 헬리케이스가 DNA 이중가닥을 풀어준다.
긴장이 만들어 지는 것을 자꾸 풀어줘야 한다. DNA gyrase가 풀어준다.
(a) 를 보면 DNA가 띄엄띄엄 합성된다.
헬리케이스가 풀고 합성하고 풀고 합성하고 하기 때문에 오카자키 조각이라고 한다.
오카자키 조각 : 연속으로 합성되지 않고 불연속으로 합성된다.
선도가닥 : 연속적으로 합성되는 가닥
지연가닥 : 불연속적으로 합성되는 가닥
★★★★★★★
DNA 중합효소 l : 지연가닥의 RNA 프라이머를 제거한다.


supercoil : 이중나선 위에 또 나선이 생기는 것
지연가닥, 선도가닥 합성 과정

RNA프라이머를 통해서 DNA가 만들어지고 있다.
지연가닥 :
베타클램프를 클램프로더가 잡아다가 RNA 프라이머가 있는 곳에 안착을 시켜줬다.
문어발처럼 생긴 클램프로더가 베타클램프를 잡기위해 모양이 변형된다.
베타클램프가 DNA를 감싸면 합성이 진행된다.
진행이 끝나면 핵심효소에 타우가 철커덕 붙어서 핵심효소를 분리하고 다시 새로운 오카자키 조각을 합성한다.
DNA 중합효소 l 이 프라이머를 제거하고 간극을 채운다.
선두가닥 : 쭉 그대로 간다.
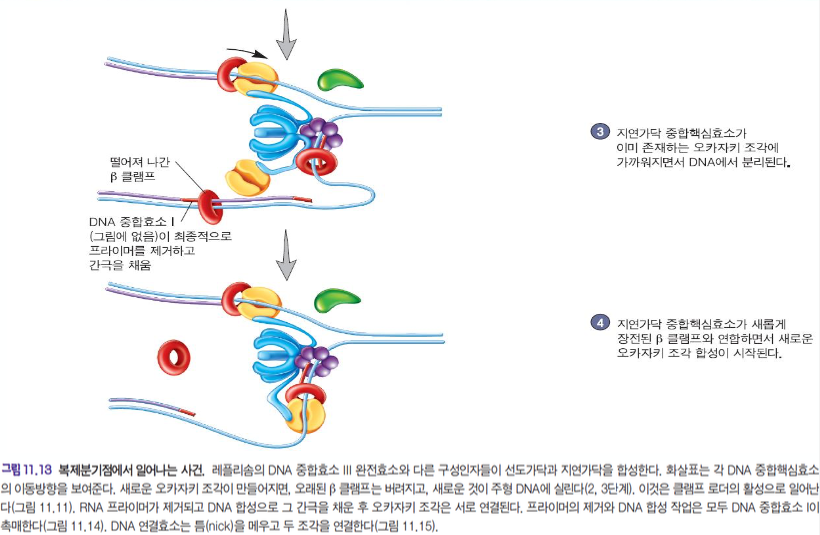

5' -> 3' 으로 가면서 오카자키 절편의 RNA 프라이머가 분해됨과 동시에 DNA 합성이 이루어진다.
이것을 DNA 중합효소 l 이 한다.
마지막 gap은 liagase가 연결한다.

DNA 중합효소 lll의 교정 활성

G가 들어가야되는데 A가 들어간다.
잘못되었으므로 이때 3'->5' 핵산말단분해 활성이 일어난다. ★★★★★★★
말단의 1개 염기를 제거한다.
복제 종결
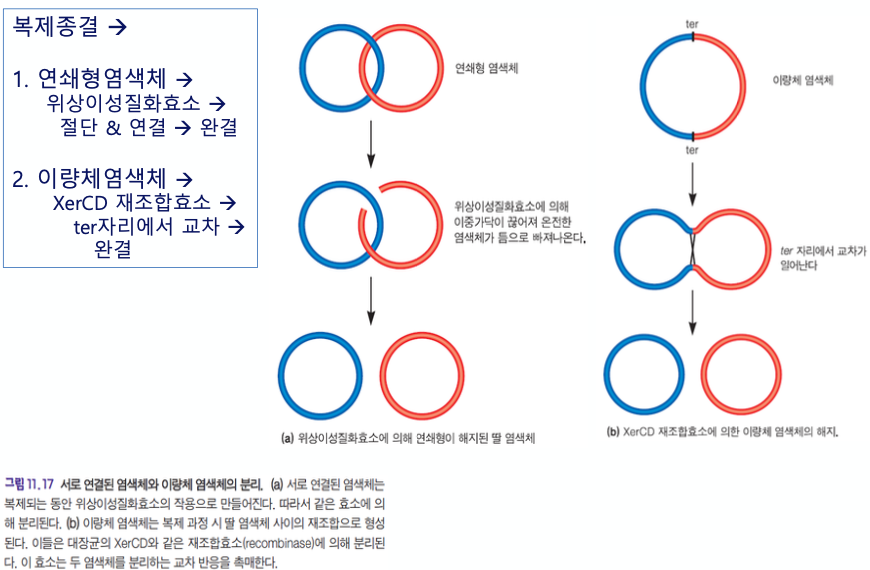
2개의 원형 DNA 분자가 만들어지다보니, 결국 고리 2개가 쇠사슬처럼 연결되어 있다.
얘네가 서로 분리되려면 decatenation이라는 작업이 필요하다.
이 때 다시 topoisomerase 친구들이 등장하게 된다.
복제가 완전히 끝나기 전에 decatenation이 일어난다면, type IA가 사용된다.
만약 복제가 끝난 후에 decatenation을 한다면, type II를 사용해서 고리를 뺀다.
재조합될 때 교차가 되면서 떨어진다.
연쇄형 염색체의 경우
위상이성질화효소가 작용한다.
이량체 염색체의 경우
XerCD 재조합효소가 작용한다. ★★★★★★★

염색체가 선형인 것처럼 보이지만 선형이 아니다.
telomere resolvase : 절단하고 연결해서 복제가 완결된다.
전사

5' -> 3' : 암호가닥
3' -> 5' : 주형가닥
주형가닥을 가져다 놓고 전사가 돼야 한다.
암호가닥이라고 하는 이유 : mRNA를 있는 그대로 읽을 수 있으므로, 암호가 되므로 암호가닥이라고 한다.
암호가닥만 읽어도 암호를 알 수 있다.
선도서열의 시작점의 +1 : 전사가 시작되는 지점 ★★★★★★★
암호화부위 : 단백질로 번역이 되는 부위, 흔히 얘기하는 유전자
후방서열
종결자
mRNA sequence에는 프로모터 서열은 없다.
선도서열, 암호화서열, 후방서열은 있다.
종결자는 없다.
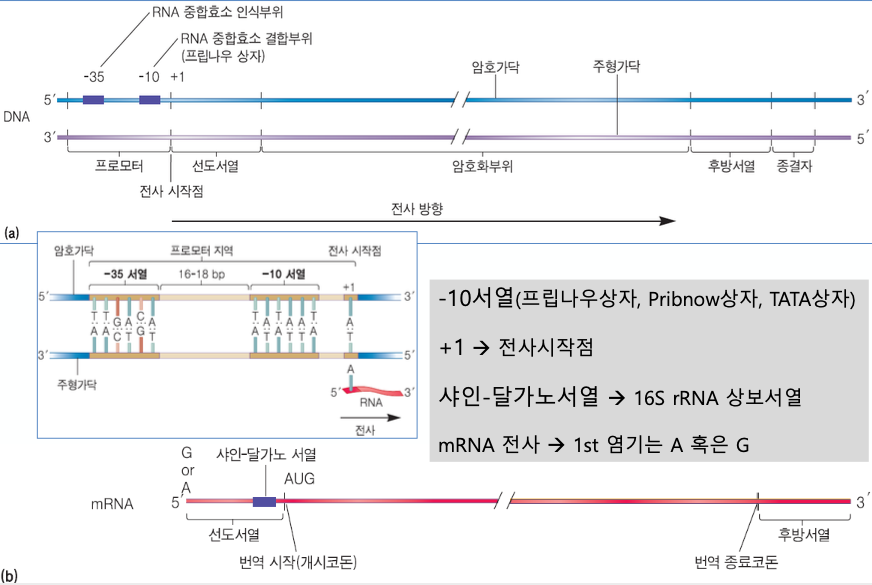
프로모터를 보면 사이에 핵심적인 부위가 있다. +35 ~ -10
-10서열 : RNA 중합효소 결합부위 : 프립나우 상자 ★★★★★★★
TATA box가 있다.
T와 A가 많다. 수소결합이 약한 부위이다.
-10 -9 .... -1 +1 >> 이런 숫자 구조
수소결합이 약한 부위가 열리게 되면서 전사의 시작이 일어나게 된다.
전사가 시작되면 +1 부터 시작된다.
선도서열이 끝나는 부위에 번역이 시작되니까 AUG가 붙는다.
번역이 시작되기 전에 샤인-달가노 서열이 있다. ★★★★★★★
샤인-달가노 서열은 16S rRNA 상보서열을 가지고 있다.
리보솜에서 번역이 될 때, 이 16S rRNA의 상보서열에 mRNA가 철커덕 붙는다.
그래야 번역이 안정적으로 될 수 있기 때문이다.
16S rRNA는 30S 에 있다. ★★★★★★★ (30S + 50S = 60S)

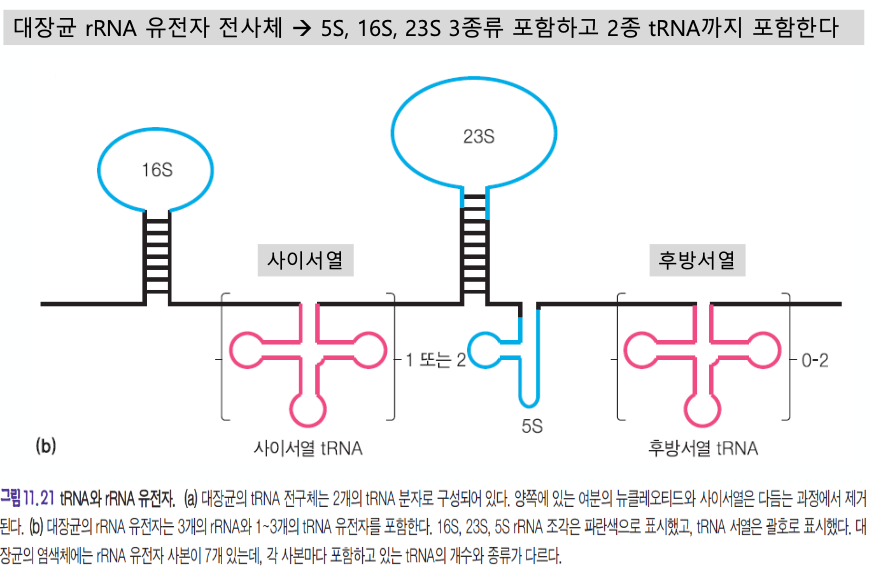
다중 시스트론 mRNA

진핵 미생물의 mRNA와 비교해보면 세균쪽의 mRNA는 유전자가 하나가 있는게 아니라 하나이상 있다.
하나의 mRNA에서 시작하고 끝나고 시작하고 끝나고 한다.
이것을 다중시스트론 mRNA 라고 한다. ★★★★★★★
RNA 중합효소는 반드시 프로모터에서 전사가 이루어진다.
그런 다음 mRNA가 전사가 된다.

'🧬 Bio > 미생물학' 카테고리의 다른 글
| 미생물학 12장 (1) - 젖당오페론, 트립토판오페론, Riboswitch (0) | 2020.06.17 |
|---|---|
| 미생물학 11장 (2) - 전사 종결, 번역 (0) | 2020.06.12 |
| 미생물학 9장 (4) - 무기영양생물의 연료공급, 광합성 (0) | 2020.06.03 |
| 미생물학 9장 (3) - 무산소호흡, 발효, 베타산화 (0) | 2020.05.30 |
| 미생물학 9장 (2) - TCA 회로, 전자전달계 (0) | 2020.05.27 |