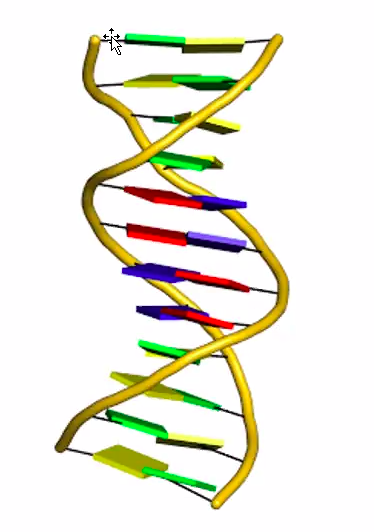

수소결합과 소수성 인력은 서로 상호 협력적이다.
이중나선구조가 유지되려면 양전기적인 요소가 있어야 이중나선 주변이 전기적으로 중성이 되고 두 사이의 거리가 가까워져서 수소결합이 이루어질 수 있다.

진핵세포의 경우 히스톤단백질이 음전하를 상쇄시켜준다.
양쪽가닥에 존재하는 염기들 사이에서 상쇄되므로 두가닥이 묶이게 된다.
안쪽으로 모여서 차곡차고 쌓여야 정렬이되고 수소결합이 가능하게 된다.
Base stacking 인력은 상호 협력적이다.
만약 수소결합을 끊으면 더 이상 마주보아야할 이유가 없으므로 구조가 무너지게 된다.
수소결합과 소수성결합은 상호 협력적이다.
또한 수소결합끼리에도 상호 협력적이고 소수성결합끼리도 상호협력적이다.
위에서 A와 T가 수소결합이 되어있으면 그 밑에 수소결합도 위가 결합되어 있으면 수소결합 하기가 쉽다.


수소결합의 중요성, 소수성 결합의 중요성을 알 수 있다.
DNA 가닥은 분리(변성)되고 재생될 수 있다.

수소결합이 깨질 수도 있다.
열을 가하면 수소결합에 관여했던 원자들의 운동에너지가 증가하기 때문이다.
변성(Denaturation) : 두 가닥이 풀어지는 것
재생(Renaturation) : 두 가닥이 다시 붙는 것
분리되어도 상보될 수 있는 짝이 있기 때문에 다시 수소결합 할 수 있다.

두가닥이 헤어졌을땐
염기가 이중나선 안쪽에 쌓여있을 때는 노출이 되어있을 때보다 덜 흡수된다.
그래서 단일가닥으로 흩어져 있을때 흡광도가 37퍼 정도 증가한다.

온도가 어느이상 올라가면 버티지 못하고 흡광도가 올라간다.
이는 염기쌍이 터진다는 것을 의미한다.
Tm = 절반정도 변성되었을 때의 온도

온도가 급작스럽게 변한다는 것은 이것을 유지하는 힘에 상호협력적이라는 것이다.

처음에는 온도를 올려도 버틴다.
절반정도 깨졌을 때 Tm값 이라고 한다.
이것은 특정 DNA의 변성온도라고 한다.
더 온도를 올리면 나머지 수소결합도 깨지게 된다.
처음에 이중결합이 깨지는 부분은 A=T 쌍이 먼저 깨지고 그 다음 깨지는 부분은 G=C 쌍이다.
수소결합이 A=T가 두개이고 G=C는 세개이기 때문이다.

GC 함량이 더 높을수록 Tm값이 더 높다.
GC 함량이 더 높을수록 더 안정하다.
같은 GC 함량이라도 양이온이 높으면 Tm 값이 높다.
이미 음전하를 충분히 가려주면 얘내들은 결속력이 있어서 잘 안 헤어진다.
양이온이 적다면 (-)척력때문에 더 잘 헤어진다.
Denaturant

수소결합을 깨는 시약
순간적으로 굉장히 짧은 시간이지만 수소결합이 깨졌다 붙였다 한다. 형광등처럼
DNA도 깨졌다 붙였다 하므로 계속 붙어있는 것처럼 보인다.
수소결합이 순간적으로 깨질 수 있기 때문에 돌연변이나 화학적변화가 일어날 수 있게 된다.
수소결합이 깨졌다가 다시 붙기 전에 작은 분자가 붙어버리면 수소결합이 떼어지게 된다.
소수성결합을 약하게 하는 시약
물과의 친화도를 증가시키면 소수성 인력이 약화된다.
왕따 당하지 않기 때문에 소수성 인력이 약화된다.
그래서 소수성 인력을 약화시키면 더 빨리 변성이 일어날 수 있다.

이중나선 입체구조를 유지시키는 힘

꼬임의 총 수 = 나선수 + 초나선 수 이다.
Lk0 = 완전히 이완됐을 때의 고리 수
대부분 진핵생물의 DNA는 선형(Linear form)이다.
그러나 원핵생물들에게선 원형(Circular form)의 DNA도 발견되는데 이것을 플라스미드(Plasmids)라고 부르고 있다.
여기서는 주로 원형 DNA. 더 정확하게 말하자면 cccDNA(Covalently colsed circular DNA)에 대해서 다루게 된다.

cccDNA는 대략 이렇게 모식화 할 수 있을 것이다. 그렇다면 이 DNA는 어떻게 전사나 복제를 하게 될 것인가?
그냥 한 군데 틈내서 양쪽으로 쭉 벌리면 되지 않을까요?
틀렸다. 이 DNA는 완전히 닫힌 형태의 나선 구조를 하고 있기 때문에 그냥 그렇게 쭉 잡아당기게 되면 나선(꼬인 것)이 전혀 풀리지 않게 된다.
cccDNA에서의 나선 수를 나타내는 것이 바로 Lk(또는 L)인데, 고리수(Linking number)라는 뜻이다.
고리수는 어떻게 알 수 있는가? 간단하다. 저 DNA의 중간을 툭 잘라서 선형 DNA로 만들고, 이중나선을 풀면서 얼마나 꼬여 있는가를 세면 된다.
그게 귀찮고 번거롭다면 저 그림에서 두 선이 겹치는 교차점 중 빨간색이 녹색(또는 녹색이 빨간색)위로 지나가는 교차점만을 모두 세면, 그것이 바로 Lk가 된다. 직접 세어보면 알 수 있을 것이다.
그런데 만약 cccDNA 이중나선이 일부 풀린 상태라면 어떻게 되는가?

가장 왼쪽은 잠시 신경을 꺼 두어도 좋다. 2형 DNA 이성질화효소(Gyrase, Type-Ⅱ DNA topoisomerase)는 초나선(Supercoil)을 만드는 효소이나, 지금 단계에선 언급할 녀석이 아니다.
내가 말하고자 하는 것은 가장 오른쪽의 녀석이다. 좁은 의미에서의 cccDNA는 저것과 같이 초꼬임이 나타나지 않은 상태를 뜻한다.
어쨌거나, 비교를 위해 왼쪽의 녀석과 비교해보면, 본래 모든 염기쌍이 결합하고 있는 상태의 Lk는 36인데 그러나 오른쪽의 녀석은 일부의 염기쌍이 풀려있어 Lk가 32다.
(Lk는 완전히 풀린 상태의 cccDNA를 기준으로 하여 센다)
즉, 나선 수가 4번 적다는 뜻이다. 총 염기쌍이 360 bp이고, 이상적인 조건 하에서 B-DNA의 1회전당 염기쌍은 10쌍이므로, 대략 40개의 염기쌍이 결합을 하지 않고 있는 상태다.
그러면 저 cccDNA는 그냥 염기쌍을 풀어놓은 상태로 존재할까? 아니다. 어떻게든 결합하려 할 것이다.
그런데 문제는, 저 상태에서 그대로 염기쌍들이 서로 결합하게 되면 본래 나선 수(Lk)는 32개 그대로인데 정상적인 상태에서 저 cccDNA가 꼬여야 할 수(Tw)는 36이기 때문에 구조가 불안정해진다.
그렇기 때문에 구조를 안정화하기 위해 cccDNA의 원형 구조 자체를 비틀려는 힘이 발생하게 되어 cccDNA 전체가 꼬이는 현상이 나타나는데, 이것이 초꼬임이다.
이거 말로 설명하려니 굉장히 어렵다. 이 설명을 듣고 이해할 수 있다고 생각되지 않는다(...)
(역학을 전공하거나 지식이 있는 사람이라면 이해할 수 있을...려나? 일단 본인부터 그런 지식이 거의 전무하다보니 설명하기가 어렵다)
다시 위의 모식도를 보자. 방금 설명을 한 것과 같이 Lk 아래의 Tw(T)와 Wr(S)가 DNA가 일부 풀려 있던 상태의 원래 cccDNA 형태와 비교하여 달라졌다는 것이 보일 것이다.
(모식도는 꽤나 예전 자료라 약어가 다르지만 최근 표기를 준수하기로 하겠다)
Tw는 말 그대로 꼬임수(Twist number)이며, Wr는 초나선수(Writhing number)이다. 꼬임수는 현재 결합한 염기쌍 수/10. 그러니까 본래 이 cccDNA가 가져야 할 나선 수이다. 위에서는 40쌍의 염기쌍이 모두 결합하였으므로 4가 늘었다.
(물론 생리적 조건에서는 /10.5를 해야 하지만 계산의 편의를 위해 이상적 조건으로 가정하였다)
물론 반대로 생각해서 Lk가 32일때 강제로 염기쌍을 30개를 더 풀어버렸다고 가정하면 Tw는 3이 줄어 29가 될 것이다.
(Lk는 변하는 값이 아니라는 점에 주의하자)
Lk와 Tw가 차이가 발생함에 따라, 그 차이만큼 초꼬임이 발생하게 되고, 따라서, Wr는 Lk-Tw로 나타낼 수 있다. Wr가 음수일 경우 음성 초꼬임(Negative supercoiling), Wr가 양수일 경우 양성 초꼬임(Positive supercoiling)이 일어난다.
그리고 이 초꼬임의 종류에는 크게 두 가지가 있는데 서로 얽힌(Interwound) 초꼬임과 토로이드 형(Toroidal) 초꼬임이 있다.
Lk(고리수) = Tw(꼬임 : 코일수) + Wr(비틀림 : 양성 초나선 수)
초꼬임
초꼬임은 단어부터 어렵다. 대체 어떤 것들인가?

대충 이런 구조이다. 토로이드형은 진핵생물의 뉴클레오솜(Nucleosome)에서 DNA가 히스톤(Histon) 단백질을 감고 있는 형태에서 나타나며, 보통 플라스미드와 같은 cccDNA에선 많은 수가 서로 얽힌 초꼬임을 갖고 있다.
저기 모식도에서 나타난 초꼬임 역시 서로 얽힌 초꼬임이다. 하지만 토로이드형이 무조건 축이 되는 다른 것들을 감고 있는 것은 아니다.
자연계의 거의 모든 생물은 음성 초꼬임을 갖고 있는데, 양성 초꼬임이 매우 안정된 형태이긴 하지만 너무 안정하기 때문에 초꼬임을 푸는데 너무 큰 에너지가 필요하기 때문이다(...)
(그렇기 때문에 펄펄 끓는 곳에서 살아가는 극호열균은 양성 초꼬임을 가지고 있어 DNA가 풀리지 않게 한다)
음성 초꼬임은 위에서도 말했다시피 나선 수(Lk)에 비해 꼬임 수(Tw)가 많을 때, 즉, 염기쌍이 일부 풀린 상태의 cccDNA에서 나타나는 구조라고 했다.
DNA는 반시계 방향(오른손 감기)으로 꼬인 이중나선이므로, 나선이 더 꼬이려는 힘이 발생하게 되면 cccDNA의 전체 구조에 반시계 방향의 힘이 작용하게 된다.
(이걸 상상하는것은 굉장히 어려울 것이다. 선형이라면 모를까 원형 DNA이므로 더 힘들다)
이 힘을 해소하기 위해서 cccDNA 전체가 초꼬임수(Wr)만큼 반시계방향(오른손 감기)으로 서로 얽히듯 꼬이게 되는데 이것이 서로 얽힌 초꼬임이다. 물론 양성 초꼬임은 DNA의 나선이 풀리려는 힘이 발생하므로 시계방향(왼손 감기)로 꼬인다.
그럼 토로이드 형은 어떤가요?
토로이드 형 초꼬임은 서로 얽힌 초꼬임과는 반대다. 즉, 음성 초꼬임에서는 시계방향(왼손 감기)으로 꼬이며, 양성 초꼬임에선 반시계방향(오른손 감기)로 꼬인다.
아니 왜요?

꼬이지 않은 부분을 생각하자. 꼬이지 않은 부분은 정상적인 나선 구조를 갖고 있어야 한다, 그렇다면 꼬이는 부분을 따로 위의 모식도처럼 떼었다고 생각하고 꼬이는 힘이 생기는 것을 보자.
당연히 반시계방향(오른손 감기)으로 힘이 작용한다. 그러면 저렇게 시계방향(왼손 감기)으로 쌓이듯이 꼬이게 된다.
(이해가 안 되면 직접 끈을 꼬아서 해 보면 된다! 쉽게 구할 수 있는 끈이 없다면 두루마리 휴지를 적당히 찢어서 꼬아도 된다)
하지만 저런 토로이드 형 초꼬임은 cccDNA에서는 무슨 단백질을 DNA가 감고 있을 때나, 축이 되는 단백질이 없다면 정말 완벽하게 원통에 코일을 감아놓은 듯한 토로이드 형이 아니라면 거의 나타나지 않는다.
관찰되는 cccDNA의 대부분의 초꼬임 형태가 서로 얽힌 형태기 때문에, 토로이드 형에 대해선 많은 이야기를 하지 않을 것이다.
위상이성질화효소 (Topoisomerase)
위상이성질화효소 1, 3 : nick을 만들어 한가닥을 끊었다가 다시 연결하여 초나선을 풀어준다 -> ATP 소모 X
위상이성질화효소 2, 4, gyrase : 두가닥을 끊었다가 다시 연결해여 초나선을 풀어준다 -> ATP 소모 O
간단히 정리하자면,
- Wr = Lk - Tw. Wr가 음수일 경우 음성 초꼬임(Negative supercoiling), Wr가 양수일 경우 양성 초꼬임(Positive supercoiling)이 나타난다.
- 초꼬임에는 서로 얽힌(Interwound) 초꼬임과 토로이드 형(Toroidal) 초꼬임이 있는데, B-DNA에서 서로 얽힌 초꼬임의 경우 음성 초꼬임은 반시계방향(오른손 감기)의 꼬임이 발생하고, 토로이드 형에서는 시계방향(왼손 감기)의 꼬임이 발생한다.
- 극히 일부의 생물체를 제외한 거의 모든 생물체는 음성 초꼬임을 주로 갖는다.
정도가 될 것이다. 3줄 요약이 가능한 것으로 포스트 하나를 장구하게 잡아먹었다(...)
어쨌거나, 여기에선 이제 저 위의 내용을 이해했다고 가정하고(...) 진행하도록 하겠다.
여기에서는 Lk0 라는 녀석과 ΔLk라는 새로 등장하게 될 텐데, ΔLk는 사실상 Wr 과 같은 뜻이다.
우선, Lk0(Lk knot)은 초꼬임이 존재하지 않고 완전히 풀린 상태(Fully relaxed form)로 존재할 수 있는 cccDNA의 Lk를 뜻한다.
말만 어렵게 해 놨지 그냥 DNA의 염기쌍 수에서 10.5로 나누면 된다(...) 즉, Lk = Tw 상태일 때의 Lk를 뜻한다.
(이중가닥 DNA는 약 10.5개의 염기당 한바퀴일 때 가장 안정한 상태이다.)
하지만 여기에서 혼동하기 쉬운 개념이 있다. Lk는 본래 나선의 꼬인 수를 나타내는 것이므로 소수가 나타날 수 없다.
즉, 완전히 이완된 상태의 cccDNA염기쌍이 100쌍인 cccDNA의 Lk와 Lk0을 계산하면 둘 다 약 9.52지만, Lk는 소수점을 버리고 9가 된다.
(10.5bp당 1회전 하는 것으로 계산하였다)
반면 Lk0은 소수로 나타날 수 있다. Tw나 Wr 역시 소수로 나타낸다. Lk만 소수로 나타나지 않는 것이다.
그렇기 때문에 사실 염기쌍의 길이가 10.5의 배수가 아닌 이상 cccDNA는 엄밀히 따져 완전히 풀린 상태를 유지할 수 없다.
비록 Wr가 1이 아니기 때문에 서로 얽히는 형태의 초꼬임이 나타나지는 않지만, 고차 구조의 뒤틀림이 발생하기 때문이다.
어쨌거나, 그렇다면 ΔLk는 어떻게 나타나는가?
ΔLk는 ΔLk = Lk - Lk0 으로 구한다. Lk0이 Lk = Tw 상태의 Lk이므로 곧 ΔLk = Lk - Tw와도 같다.
Lk - Tw 는 무엇인가? 바로 위에 있다. Wr이다. 즉, ΔLk = Wr 이라는 공식이 간단히 성립한다.
그런데 왜 이걸 굳이 또 만들었느냐. 모르겠다 내가 만든게 아니다(...)
어쨌든, 이걸 이용하여 우리는 DNA의 초나선 꼬임 밀도(Superhelical density) σ를 구할 수 있다. 다음과 같다.
σ = ΔLk / Lk0. Lk0이 작을수록, Wr가 클 수록 초나선 꼬임의 밀도가 크다는 뜻이다.
그리고 같은 크기의 DNA라면 σ 가 클 수록, 전기영동에서 DNA가 이동하는 속도가 더 빠르다. 훨씬 조밀한 구조니까 상식적으로 당연하다 생각할 수 있겠다.
(가장 느린 것은 단일 사슬 DNA이며, 그 다음으로 완전히 풀린 상태의 cccDNA, 그 다음으로 선형 DNA 순이다)
그런데 실험실에서 DNA를 전기영동을 시킬 때 EtBr을 이용하여 염색을 하는 경우가 많다. 여기에 포함된 에티듐이라는 양이온은 DNA의 사이에 끼어들어가 1회전 당 염기쌍 수를 늘인다.
무슨 뜻인가? 이상적 조건 하에서 B-DNA의 1회전 당 염기쌍 수는 평균 10bp다. 그러나 에티듐 이온이 하나 첨가되면, 에티듐 이온이 첨가된 곳의 DNA 회전 각도를 36˚에서 10˚로 감소시킨다.
(생리적 조건과 이상적 조건은 조금 다르지만, 생리적 조건은 딱 떨어지지 않아 계산이 불편하여 편의상 이상적 조건을 사용하였다)
즉, 모든 염기쌍 사이에 에티듐 이온이 끼어들어가게 되면 이상적 조건 하에서 B-DNA의 1회전 당 염기쌍 수는 평균 10bp에서 평균 36bp로 증가하게 된다.
무슨 뜻인가?
Tw가 줄어든다는 뜻이다. Tw는 염기쌍 수/10 이었으나 에티듐이 추가됨에 따라 염기쌍 수/36 까지도 변할 수 있다는 뜻이다.
Tw가 줄어들게 되면 Lk = Tw + Wr 이므로 Wr는 Lk에 맞춰서 증가해야 할 것이다. 이 결과, 본래 음성 초꼬임을 갖던 DNA의 초꼬임 밀도가 낮아지거나, 또는 아예 양성 초꼬임으로 변할 수도 있게 된다.
만약에 완전히 풀린 상태의 cccDNA라면 Wr는 0이다. 그러나 여기에 에티듐 이온을 첨가하면 Wr는 증가한다. 그러면 양성 초꼬임이 나타나게 될 것이다.
전기영동이 끝난 후에 염색을 할 경우에는 문제가 되지 않으나, 실험의 편의성을 위해 겔에 미리 EtBr을 첨가하게 되면 에티듐 이온에 의해 초꼬임 밀도가 바뀌어 제대로 된 전기영동 결과를 얻을 수가 없게 된다.
자, 그러면 이제 이런 위상이성질체에 대해 알았으니 이에 관여하는 효소에 대해 알아볼 차례이다.
많은 박테리아 DNA들이 음성 초꼬임의 형태를 하고 있다. 하지만 이 상태로는 DNA의 전사나 복제가 불가능할 것이다. 즉, 초꼬임을 풀어주어야만 한다는 말이 된다.
그리고 전사나 복제가 끝나면 다시 초꼬임의 형태로 되돌려 놓아야 한다. 여기에 관여하는 효소는 크게 두 종류의 효소가 있다.
하나는 제 1형 DNA 위상이성질화효소(DNA topoisomerase type-Ⅰ)이며, 다른 하나는 제 2형 DNA 위상이성질화효소(DNA topoisome-rase type-Ⅱ)이다. 2형의 경우 음성 초꼬임을 유도하는 녀석들을 특별히 자이라아제(Gyrase)로 부르기도 한다.
1형의 경우는 DNA 단일 사슬에 결합하여 Lk를 1만큼 변화시킨다. 어떻게 하는가?
간단하다. DNA 단일 사슬에 결합하여 그 사슬을 끊고, 반대편 사슬 뒤로 넘겨서 결합시킨다. 그러면 결과적으로 꼬임이 하나 더 늘어나거나 풀리게 되어 Lk가 1만큼 변하게 된다.
2형의 경우는 DNA 이중 사슬에 결합하여 이중 사슬 전체를 끊고, 다른 사슬 뒤로 넘겨서 결합시킨다. 이 경우엔 Lk가 2만큼 변하게 된다.
(직접 끈을 꼬아서 해보면 이해가 빠를 것이다)

이들의 범용성은 무궁무진하여, 이런 다양한 꼬임들을 모두 풀어낼 수 있다. 물론 다시 만들어 내는 것도 가능하다.
그러나 이들은 단지 매듭을 푸는데만 이용되는 것은 아니다. DNA가 복제될 때를 생각해 보자.
DNA가 복제되기 위해선 DNA 이중 나선이 풀려야 한다. 그러나 DNA 단일 사슬 하나씩을 양 손에 잡고 쭉 찢는다고 상상해보라. 당연히 꼬여 있으니 풀리면서 빙글빙글 돌아가게 될 것이다.

DNA가 복제되는 곳 앞에서 계속 이중나선을 풀어주면서 복제가 원활하게 되도록 하는 것 역시 이들이 하는 역할이다.
또 한, 복제된 딸 DNA 역시 서로 꼬이거나 할 수 있는데, 이 때에도 2형 이성질화 효소가 꼬인 것을 풀어주게 된다.
그렇다면 이들은 어떻게 DNA의 결합을 끊고 재결합시키는가?
1형의 경우 ATP를 필요로 하지 않는다. 그러나 2형의 경우 좀 더 큰 일(...)을 하기 때문에 ATP 2분자를 필요로 한다. DNA 이중 사슬을 아예 끊고 다시 연결하는 것이 쉬운 일은 아닐 것이다.
이들은 공통적으로 인산-티로신 공유결합을 이용하여 DNA를 절단하게 되는데, 활성화된 티로신 잔기가 DNA 뼈대의 인산 결합을 끊고 자신과 결합시킴으로써 일어난다.
물론 반대편에서도 아예 끊어져서 도망가지 않게 효소와 DNA 말단의 결합이 일어나게 되어 효소가 일종의 다리 역할을 하며 DNA를 양 손으로 잡고 있는 듯한 형태가 된다.
이 후, 효소의 구조가 크게 변해 반대편 사슬을 넘기거나, 혹은 사슬 전체를 돌려 DNA의 회전수(Lk)를 변화시켜 다시 결합시킨다.
'🧬 Bio > 분자생물학' 카테고리의 다른 글
| 분자생물학 9장 (2) - 복제분기점, Primase, Helicase, DNA polymerase (6) | 2020.10.10 |
|---|---|
| 분자생물학 9장 (1) - DNA 복제 (0) | 2020.10.04 |
| 분자생물학 8장 (2) (0) | 2020.09.25 |
| 분자생물학 8장 - 유전체 구조, 염색질 그리고 뉴클레오솜 (0) | 2020.09.25 |
| 분자생물학 4장 - DNA 입체구조 (1) (0) | 2020.09.08 |