mRNA - Pol ll
우선은 가장 잘 알려진 mRNA의 전사에 대해 살펴보도록 하자. mRNA는 누가 담당한다? 제 2형 RNA 중합효소(Pol ll)가 담당한다.
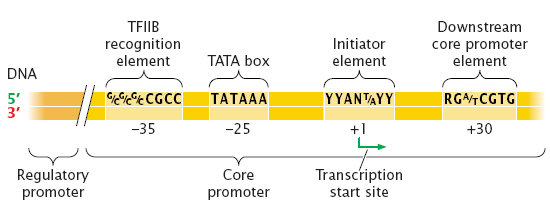
진핵생물의 프로모터(Promoter)로 가장 잘 알려진 것은 TATA 박스(TATA box)라 불리는 염기 서열 부위다.
그러나 RNA 중합효소가 스스로 프로모터를 인식하여 결합하지는 못한다. DNA 복제와 비슷하게 RNA 전사 과정에서도 프로모터에 결합하는 여러 전사 조절 단백질들이 복합체를 형성해야 한다.
시험관 조건(In vitro)에서는 DNA 주형에 RNA 중합효소와 프로모터에 관여하는 몇몇 보편 전사인자(GTF, General transcription factor)를 넣어주게 되면 전사가 일어난다.
그러나 실제 생리적 조건(In vivo)에서는 DNA가 뉴클레오솜과 염색질(Chromatin)의 형태를 하고 있다.
따라서 보편 전사인자 외에도 매개복합체(Mediatior complex)나 여러 DNA 결합 단백질(DNA binding proteins), 히스톤 샤페론(Histon chaperon) 등이 더 필요하다.
Promoter
다음으로, 진핵생물의 프로모터 부위는 어떻게 되어 있는가?
프로모터는 정확하게 말하면 모든 전사 개시 및 조절 인자들이 결합하는 염기서열을 뜻한다. 즉, 전사 개시 뿐만 아니라 조절에 영향을 주는 염기서열 역시 프로모터라 볼 수 있다.
RNA 중합효소가 전사를 개시하기 위해 필요한 최소한의 염기서열을 핵심 프로모터(Core promoter)라 부르는데, 핵심 프로모터는 약 40~60b 길이로 구성되어 있다.
역시 가장 잘 알려져 있는 것은 상류 25b 정도에 위치한 TATA 박스지만 TATA 박스로만 프로모터가 구성되진 않는다.
(상류란 전사가 시작되는 전사 개시점의 앞부분을 뜻한다. 반대로 전사 개시점의 뒷부분은 하류라 부른다)
핵심 프로모터에는 대략 다음과 같은 4개의 구성요소가 있다. 그러나 일반적으로 이 4가지가 모두 포함된 경우는 별로 없으며 2~3개 정도의 요소가 모여 핵심 프로모터를 형성한다.
(즉, 어떤 유전자의 프로모터에서는 TATA 박스가 없을 수도 있다는 뜻이다)

BRE : TF ll B recognition element
우선, 가장 상류에 위치한 BRE(TFllB recognition element)는 차후 언급하겠지만 보편 전사인자 중 하나에 속하는 TFllB 단백질이 결합하는 부분이다.
TATA box
TATA 박스는 마찬가지로 보편 전사인자에 속하는 TFllD가 인지하여 결합하는 부분으로, TFllD가 TATA 박스에 결합하는 것으로 개시전 복합체(Pre-initiation complex)의 구성이 일어난다.
Initiator element (개시요소)
개시 요소(Inr, Initiator element)는 TATA 박스와 기능적으로 유사하다. TATA 박스가 없고 개시 요소만 있을 경우 TFllD가 개시 요소에 결합하여 전사를 개시한다.
Downstream Promoter Element (DPE)
하류 프로모터 요소(DPE, Downsteram promoter element)역시 RNA의 전사를 개시할 수 있으며 주로 개시 요소와 함께 조절한다. 여기에서는 자세한 설명은 하지 않을 것이다.
Regulatory sequence (조절 서열)
그러면 핵심 프로모터 외에 다른 프로모터 요소에는 어떤 것들이 있는가? 조절 서열(Regulatory sequence)라 불리는 요소들이 있다.
조절 서열은 주로 핵심 프로모터보다 훨씬 상류에 위치하는 경우가 많으며, 어떤 것들은 수백 kb 떨어진 위치에 존재하기도 한다.
극단적으로는 아예 다른 염색체에 위치하기도 하는데, 황당한 소리로 들리겠지만 DNA는 항상 3차적으로 꼬이고 접혀진 구조를 하고 있다.
따라서 조절 서열과 핵심 프로모터 부분이 서로 상호작용할 수 있을 정도로 충분히 가까워질 수 있으므로 불가능한 것은 아니다.
이러한 조절 서열에는 프로모터 근접요소(Promoter proximal element), 상류 활성서열(UAS, Upstream activator sequences), 증폭인자(Enhancer), 침묵인자(Sliencer), 경계 요소(Boundary element), 절연인자(Insulator) 등이 있다.
이들은 주로 직접적으로 전사를 개시하거나 진행되는데 필요한 서열은 아니지만 전사를 촉진하거나 억제하는 역할을 담당한다. 말 그대로 전사 수준을 조절하는 역할이다.

그러면 이제 가장 보편적인 전사 기작을 살펴보도록 하자. 우선은 Pol ll에 의해 TATA 박스로부터 전사가 어떻게 개시되는지 보자.

TFllD
먼저, TATA 박스에 TFllD가 결합한다. TFllD는 여러 서브유닛으로 구성된 복합체인데, 그 중 TBP(TATA binding protein)라는 서브유닛에 의해 TATA 박스를 인식하여 결합할 수 있다.
TFllD에는 TBP 외에 여러 종류의 TAF(TBP associated factors)가 있는데, 어떤 TAF는 TBP와 TATA 박스 사이의 결합을 도와주는 역할을 한다.
또 다른 종류는 위에서 언급했던 개시 인자(Inr)나 하류 프로모터 요소(DPE, DCE 등)에 결합하여 전사를 개시하기도 한다.
즉, 우선 전사가 개시되기 위해선 TFllD가 핵심 프로모터에 결합해야만 한다.
특이한 점은 TBP는 TATA 박스의 넓은 홈(Major groove)이 아니라 좁은 홈(Minor groove)을 인식한다는 점이 특이하다. 대부분의 DNA 결합 단백질은 DNA의 넓은 홈이 갖는 화학 정보를 인식한다는 점을 기억하는가?
어쨌거나 TBP(TFllD)가 TATA 박스에 결합하고 나면 DNA의 구조가 변형되는데, TATA 박스 부위에서 약 80도 정도로 DNA 사슬이 꺾여 TBP 결합부위의 작은 홈이 완전히 노출되게 된다.
TFllA
그 후 뒤이어 TFllA와 TFllB가 따라 들어오게 되는데, TFllA는 TBP와 TATA 박스 사이의 결합을 더욱 안정화하고 부자연스럽게 꺾여진 DNA의 구조를 유지하는 기능을 하는 것으로 여겨진다.
TFllB
TFllB 또한 TFllD와 서로 상호작용을 하게 되는데, 재미있는 것은 이 단백질은 DNA가 꺾인 부분에 결합한다.
그래서 TATA 박스의 상류에 위치한 BRE 서열의 넓은 홈과 TATA 박스 하류에 위치한 염기서열의 좁은 홈과도 서로 상호작용을 한다는 점이다.
단순히 설명으로는 이해가 잘 안 되겠지만 모식도를 보면 이해가 수월할 것이다. 대략 이러한 형태를 하고 있다.

잘 보면 이들의 결합이 비대칭적으로 이루어져 있다는 것이 보이는데, 이로 인해 RNA의 전사는 하류 방향으로만 일어나게 된다.
또한 TFllB는 Pol ll가 결합할 수 있도록 하는 역할을 하여 결과적으로 TBP-TFllB-Pol ll의 결합을 형성한다.
그러고 나면 드디어 RNA 중합효소(Pol ll)가 들어오게 되는데, Pol ll 혼자 들어오는 것이 아니라 TFllF라는 녀석이 결합한 상태로 같이 들어온다.
TFllF
TFllF는 TBP-TFllB-Pol ll의 결합을 안정화시키고, TFllE와 TFllH가 들어오기 위해 필요하다.
최종적으로 TFllE와 TFllH가 순서대로 들어옴에 따라 개시 전 복합체가 형성된다. 하지만 아직까지도 전사는 일어나지 않는다.
TFllE
TFllE는 TFllH의 결합을 안정화시키고 TFllH의 기능을 적절히 조절한다.
전사 개시에 있어 중요한 것은 TFllH인데, 이 녀석은 대략 10개의 서브유닛으로 구성되어 다른 보편 전사인자들에 비해 월등히 크다.
서브유닛이 많다는 것은 무엇을 뜻하나? 당연히 탑재하고 있는 기능의 수가 많다는 것을 뜻한다.
TF ll H
TFllH는 헬리카아제(Helicase)와 ATPase 활성을 갖고 있어 프로모터 부위의 DNA를 풀어 전사를 준비한다.
ATPase 활성을 갖는 서브유닛들은 또 한 다른 인자들과 함께 잘못된 리보뉴클레오티드가 짝지워졌을 때 이것을 수선하는 역할도 한다.
그러나 이것 외에 TFllH는 인산화효소(Kinase) 활성 역시 갖고 있어 Pol ll의 C 말단 꼬리(C-terminal domain tail)을 인산화한다.
RNA 중합효소의 C 말단 꼬리를 인산화하면 어떤 일이 일어나나요?
단순히 프로모터 영역에서 개시전 복합체가 제대로 구성되었다 하더라도 RNA 전사가 곧바로 일어나는 것은 아니다.
개시전 복합체가 형성되고 나면, Pol ll는 프로모터에 고정된 상태에서 덜거덕거리며 반복적으로 짧은 RNA 단편들(대략 10b 정도)을 만들다가 전사에 실패하는 모습을 보인다.
이것은 Pol ll가 프로모터에 그대로 고정된 상태에서 전사를 시작하려다보니 Pol ll가 앞으로 전진하는 것이 아니라 DNA를 뒤로 끌어오는 방식으로 전사를 하다가 결국 Pol ll에서 DNA가 튕겨져 나가며 전사에 실패하는 것으로 여겨진다.
어쨌거나 전사가 정상적으로 진행되기 위해서 필요한 것이 Pol ll의 프로모터 탈출(Promoter escape)이며 이것에 필요한 것이 Pol ll의 C 말단 꼬리의 인산화다.
Pol ll의 C 말단 꼬리는 7개의 염기(Tyr-Ser-Pro-Thr-Ser-Pro-Ser)가 여러번 반복되는 구조로 되어 있으며 TFllH의 인산화효소 활성에 의해 염기들이 인산화됨에 따라 프로모터와 대부분의 보편 전사인자들에게서 Pol ll가 탈출하여 전사가 시작되게 된다.
또 한, Pol ll의 C 말단 꼬리의 어느 부위에 인산화가 일어나느냐에 따라 전사 과정에서 필요한 여러 단계들에 필요한 효소들의 활성을 조절할 수 있다.
무슨 뜻인가? 완성된 mRNA는 5' 말단에 캡(Cap)이 씌워져 있고, 이어맞추기(Splicing)을 통해 인트론(Intron)을 모두 제거한 상태이며, 3' 말단에는 폴리아데닐화(Polyadenylation)가 되어있어야 한다.
하지만 이 단계는 전사가 모두 끝난 상태의 pre-mRNA에서 일어나는 단계가 아니라 전사 도중에 일어나게 된다.
즉, 5' 말단의 캡 형성은 전사가 시작된 직후에 일어나게 되며, 이어맞추기는 엑손(Exon)에서 다음 엑손까지 중합이 완료되면 그 사이의 인트론을 제거하며 일어난다.
A염기 반복 역시 전사가 종결되기 바로 전에 pre-mRNA의 3' 말단에서 일어나야 하는 단계다.
이 단계를 조절하는 방법이 Pol ll의 C 말단 꼬리의 특정 부분에 인산화를 통해 일종의 마커로써 활용해, 미리 효소를 준비시켜 두었다가 필요할 때 바로바로 사용하는 것이다.
RNA의 전사는 전 포스트에서 살펴보았던 보편 전사인자(GTF, General transcription factors)와 RNA 중합효소(RNA Polymerase)만 존재해도 충분히 일어날 수 있다.
하지만 RNA 전사를 조절하기 위한 여러 조절 서열(Regulatory sequence)들이 있던 것을 기억하는가?
이러한 조절 서열들은 어떤 방식으로 전사를 조절할까? 이들 역시 DNA 결합 단백질들을 이용한다. 뻔하다면 뻔하다.
이것을 총괄하는 녀석이 매개복합체(Mediator complex)라는 굉장히 거대한 단백질이다.

위는 효모의 매개복합체이고, 아래는 사람의 매개복합체다.
매개복합체는 아무리 작아도 20개 이상의 서브유닛으로 구성되어 있는 복합체인데, 각각의 서브유닛은 종마다도 다르며, 하나의 개체에서도 세포의 기능에 따라 구성이 달라진다.
서브유닛이 많다는 것은 무슨 뜻이다? 기능이 다양하다. 혹은 상호작용 할 수 있는 기질이 풍부하다는 뜻이다.
매개복합체는 한쪽 면에서는 개시전 복합체(Pre-initiation complex)를 형성하고 있는 RNA 중합효소의 C 말단 꼬리와 결합하고, 다른 면으로는 조절 서열에 결합한 활성인자(Activator)와 결합한다.
각각의 서브유닛은 상호작용할 수 있는 활성인자가 모두 다르다. 그렇기 때문에 매개복합체의 다양성은 매우 클 수 밖에 없다.
그러나 Med17 이라는 서브유닛은 모든 진핵세포에게 있어 공통적으로 필수적인 서브유닛으로 여겨진다.
어쨌거나 매개복합체는 활성인자와 상호작용을 통해 어떤 역할을 하는가? 말 그대로 조절하는 역할을 한다.
증폭인자(Enhancer)와 같은 서열에 결합한 활성인자와 상호작용하게 될 경우 전사가 촉진될 것이고, 억제인자(Sliencers)와 같은 서열에 결합한 활성인자와 상호작용할 경우 전사가 억제될 것이다.

위 모식도에서, 매개복합체에 결합하고 있는 청색, 녹색, 황색의 오뚜기 같이 생긴 녀석들이 활성인자가 되며, 활성인자가 결합한 DNA 위치가 조절 서열들이다.
DNA의 조절 서열은 앞 포스트에서 언급했지만 프로모터(Promoter) 부위로부터 수십 kb에서 심한 경우는 수백 kb 떨어져 있기도 하고 극단적으로는 아예 다른 염색체에 존재하기도 한다.
하지만 위 모식도와 같이 DNA는 항상 3차원적으로 꼬이고 접혀 있기 때문에 어떻게든 가까이 위치할 수만 있다면 큰 문제는 되지 않는다.
물론 접근성의 문제가 있어 그 효과가 차이가 날 수는 있을 것이다.
만약 전사를 촉진하는 서열이 프로모터 바로 근처에 있어 쉽게 상호작용할 수 있는 것에 반해, 전사를 억제하는 서열은 멀리 떨어져 있어 상호작용하기가 상대적으로 곤란하다면 어떠한가?
그 유전자는 주로 전사가 촉진되는 방향으로 일어날 것이다. 그 정도의 차이다.
세포의 기능에 의해 발현되는 유전자의 종류도 다를 것이고, 따라서 유전자의 발현을 조절하는 매개복합체의 구성 역시 달라질 수 밖에 없다.
실제로, 인간의 발생 과정에서도 발생 단계에 따라 세포가 분화됨에 따라 각기 다른 매개복합체를 갖는 것으로 알려져 있다.
CTD
어쨌거나, 매개복합체는 이 뿐만 아니라 TFllH에 존재하는 CTD 인산화효소(C-terminal domain kinase)를 조절하여 전사가 개시되도록 도와주는 역할을 하는 것으로도 알려져 있다.
그리하여 RNA 중합효소 여기서는 Pol ll의 C 말단 꼬리의 5번 세린이 인산화되면, 드디어 프로모터를 탈출하여 전사가 개시된다.
(CTD는 Tyr-Ser-Pro-Thr-Ser-Pro-Ser의 서열이 반복적으로 진행된다)
Elongation
전사가 개시되면, 곧바로 신장(Elongation) 단계로 이행된다. 이 과정에서는 Pol ll에서 보편 전사인자들이나 매개복합체들은 거의 다 떨어져 나가고 새로운 단백질들이 관여한다.
이것들을 신장 인자(Elongation factors)라 부르는데, 신장 인자 외에도 중합된 RNA를 가공하기 위한 인자(RNA processing factors)들이 결합하게 된다.
재미있게도, 이들은 Pol ll의 C 말단 꼬리에 일어난 인산화를 마커(Marker)로 삼아 모이고, 결합한다.
구조적으로 C 말단 꼬리는 Pol ll에서 RNA가 중합되어 빠져나오는 통로와 가깝게 위치하며, 대략 800A 정도의 길이를 하고 있다.
이것은 무엇을 뜻하는가? C 말단 꼬리에 미리 RNA 가공 인자들이 결합해 있다가 RNA가 나오면 곧바로 가공할 수 있다는 뜻이다.
신장 인자에는 다양한 단백질이 있는데, 그 중 P-TEFb는 전사가 개시되고 나면 Pol ll에 결합하여 C 말단 꼬리를 이루는 반복서열 중 2번 세린을 인산화시켜 RNA가 신장되도록 한다.
또 한, P-TEFb는 다른 신장 인자인 hSPT5라는 단백질을 인산화시켜 활성화시키고, 또 다른 신장 인자인 TAT-SF1도 P-TEFb에 의해 활성이 촉진된다.
TFllS
한편, 전사를 촉진하는 전사인자로는 ELL계 신장 인자들과 TFllS와 같은 인자들이 있다. 이 둘은 공통적으로 Pol ll의 진행성을 향상시키는 역할을 한다.
DNA 중합효소와는 달리, RNA 중합효소는 중간중간 특정 염기서열에서 종종 멈추거나, 아예 떨어져 나가버리는 경우가 가끔 존재한다.
왜 그런가요? 모르겠다. 어쨌거나 이런 문제를 해결하는 신장 인자들이 위와 같은 신장 인자들의 주 목적이다.
TFllS는 다른 한편으로는 RNA 수선(RNA proofreading)을 도와주는 역할도 담당한다.
Pol ll 역시 DNA 중합효소와 비슷하게 RNA 가수분해효소 활성을 갖고 있어, 잘못 들어간 염기가 있을 경우 TFllS가 RNA 가수분해효소의 활성을 촉진하여 RNA를 수선하도록 한다.
히스톤
하지만 DNA의 복제에서도 이미 다룬 내용이나 RNA의 전사에서도 마찬가지의 문제가 존재한다. 무엇인가?
전사를 진행하다가 만나게 되는 히스톤 단백질을 적절하게 처리를 해 주어야 한다.
히스톤 단백질들은 전사를 굉장히 크게 방해하는 요소다. 생각해보면 당연하다. 따라서 FACT와 같은 몇몇 히스톤 샤페론(Histon charperone)들이 필요하다.
(어디서 본 것 같은 기억이 난다면, DNA 복제 포스트 맨 아래에 보면 찾을 수 있을 것이다)
말이 나온 김에 FACT에 대해 간단히 살펴보자. SSRP1과 Spt16이라 불리는 두 서브유닛으로 구성되어 있는데 이 서브유닛들이 중심 히스톤(Core histon)과 상호작용한다.

이것은 FACT가 작동하는 기전에 대한 두 가지 모델이다. 물론 이게 중요한 것은 아니다. 공통적으로 마지막 단계에서 Dimer(H2A·H2B 2량체)가 제거되는 것을 보라.
중심 히스톤의 구조는 어떻게 되어있다? H3·H4 4량체와 H2A·H2B 2량체 2개로 이루어져 있다.
이 중 H3·H4 4량체에는 SSRP1과 결합을 하고, H2A·H2B 2량체는 Spt16과 결합한다. 그런 후에 H2A·H2B 2량체 중 하나가 제거된다.
그러면 뉴클레오솜의 구조가 무너져 전사가 계속 진행될 수 있다. 아니 뉴클레오솜이 완전히 해체된 것이 아니잖아요? 아직 여기에 대한 자세한 원리는 많이 밝혀지지 않아서 잘 모른다.
어쨌거나, RNA 중합효소가 지나가고 나면 FACT는 다시 제거했던 H2A·H2B 2량체를 중심 히스톤에 끼워 넣어 뉴클레오솜을 조립한다.
이 기전은 RNA 중합효소의 앞 뒤에서 즉각적으로 일어나는 반응이다. 이 방법을 통해 전사는 염색질의 본래 상태를 유지하면서도 계속 진행될 수 있다.
(물론 30nm 섬유 구조는 풀려야겠지만 말이다)
RNA의 신장(Elongation) 단계까지 했다. 그러면 이제 전사의 종결(Termination)을 하기 전에 RNA 가공(RNA processing)에 대해 알아보도록 하자.
RNA, 여기에서는 mRNA가 정상적으로 작동하기 위해 필요한 가공은 다음과 같은 세 종류가 필요하다.
우선 5' 말단의 캡 형성(5' Capping), 이어맞추기(Splicing), 3' 말단의 폴리아데닐화(3' Polyadenylation).
각 단계는 재미있게도 RNA의 신장과 함께 일어난다. 신장 인자(Elongation factors)와 RNA 가공인자(RNA processing factors)들은 서로 중복되는 것이 많다는 점이 이것을 증명한다.
앞 포스트에서도 언급했던 신장 인자인 hSPT5는 막 전사가 개시되어 C 말단 꼬리의 5번 세린에 인산화가 되어있는 Pol ll의 C 말단 꼬리로 5' 캡 형성 효소(5' Capping enzyme)들을 모으고 활성을 촉진시킨다.
무슨 뜻인가? 언제 캡 형성을 하는가?
5' Capping
전사가 개시됨과 동시에 빠져나오는 mRNA의 5' 말단에 즉시 캡을 형성한다는 뜻이다. 상당히 효율적인 방식이라 할 수 있다.
캡 형성 단계는 다음과 같이 일어난다. 캡은 메틸화된 구아닌이 5' 말단에 비정상적으로 결합하는 것을 뜻하는데, 아래를 보자.

아니, 이게 무슨 소리야. 천천히 보자. 맨 위쪽은 가공되지 않은 mRNA의 '5 말단을 나타낸다. N은 염기, p는 인산이다.
(N은 무슨 염기인가요? A, G, T, U 뭐든 될 수 있다는 뜻이다)
ppp니까 삼인산기다. 다음 단계를 보면, RNA 삼인산 가수분해효소에 의해 5' 말단의 인산 하나가 떨어져나간다.
다음 단계는 구아닐릴 전이효소(Guanylyltransferase)에 의해 구아닌이 첨가되는 단계이다. 이 구아닌은 메틸화되지 않은 정상적인 구아닌이다.
이 단계에서 구아닌에서 피로인산(Pyrophosphate)이 빠져나오며 구아닌과 5' 염기 사이에 5'-5' 삼인산 결합이 일어난다. 보통 염기가 첨가되는 방향과는 반대다.
그러고 나면 이제 메틸화 전이효소(Metyltransferase)에 의해 구아닌의 2번 탄소에 메틸기가 추가되어 메틸 구아닌이 된다.
때로는 다음 염기에서도 메틸화가 일어나는 경우가 있으나, 모든 경우에서 나타나는 것은 아니다.
캡 형성 효소는 Pol ll의 5번 세린이 인산화된 C 말단 꼬리에 결합하고 있다가 2번 세린이 인산화 되기 위해 5번 세린에서 탈인산화가 일어날 경우에만 꼬리에서 떨어져 나와 자기 할 일을 한다.
그래서 캡 형성은 Pol ll가 20~60b의 리보뉴클리오티드 정도 중합한 초기에 일어나게 되는 것이다.
(C 말단 꼬리는 전사가 개시되면 곧바로 신장 인자인 P-TEPb에 의해 5번 세린의 탈인산화와 2번 세린의 인산화가 일어난다)
Pol ll의 C 말단 꼬리의 2번 세린이 인산화가 일어남과 동시에 본격적인 RNA의 신장이 일어난다. 그리고 그와 동시에 RNA 이어맞추기에 필요한 기구들이 결합한다.
RNA 이어맞추기는 굉장히 복잡한 기전이므로, 여기서는 생략하도록 하겠다. 곧 다루게 될 것이다.
마지막 단계인 3' 말단 폴리아데닐화는 전사의 종결 단계에서 일어나게 되며, 마찬가지로 이에 관련된 효소들이 C 말단 꼬리에 결합하고 있다가 특정 DNA 서열을 Pol ll가 지나게 되면 활성화된다.
이 단계에서는 RNA 절단, 3' 말단 폴리아데닐화, RNA 가수분해효소에 의해 중합효소에 남아 있는 RNA 분해, 전사 종결이 차례대로 일어난다.

우선, 3' 말단 폴리아데닐화 과정에 대해 살펴보도록 하자. Pol ll가 유전자의 끝 부분에 있는 특정 신호 서열을 중합하면 이것을 인식하여 CstF, CPSF이라는 두 효소가 C 말단 꼬리에서 분리되여 RNA에 결합한다.
(CstF는 Clevage stimulation factor 이며, CPSF는 Clevage and polyadenylation specifiuity factor의 준말이다)
그러면 CstF에 의해 RNA의 절단이 일어난다. 임무를 마친 CstF는 떨어져 나가고, 남아있는 CPSF를 인식하여 PAP(Poly-A polymerase)이 RNA의 3' 말단에 결합하여 아데노신을 반복적으로 첨가한다.
이 과정에서 첨가하는 방식은 여타 중합효소들이 3'-5' 결합으로 염기를 첨가하는 방식과 완전히 동일하지만, 특이하게도 주형이 필요하지 않다. 그야 아데노신만 무진장 달아대니까...
이 폴리아데닌 사슬의 길이는 대략 200b 이상의 제법 긴 사슬이다. 이 사슬의 길이는 항상 동일한 것이 아닌데, 아직까지 명확하게 폴리아데닌 사슬의 길이가 어떻게 정해지는지 밝혀진 바는 없다.
그러나 신장된 폴리아데닌 사슬에 여러 폴리아데닌 결합 단백질(Poly-A binding proteins)이 결합함에 따라 이 길이가 조절되는 것으로 여겨진다.
3' 말단 폴리아데닐화는 오직 mRNA에서만 나타나는 현상이다. 그런데 대체 이것은 왜 일어나는 것인가?
이것은 리보솜에서 RNA를 번역할 때 좀 더 효율적으로 하기 위한 것으로 여겨지는데 5' 말단의 캡과 3' 말단의 폴리아데닐 사슬이 서로 상호작용을 할 수 있다.
그러면 mRNA는 고리를 이루게 되는데, 그러면 어떻게 되는가? 리보솜이 mRNA에서 떨어지지 않고 mRNA를 빙글빙글 돌려가며 반복적으로 번역을 할 수 있게 될 것이다.

위에는 elF나 PABPI같은 것은 연결 단백질들이다. 아래에는 리보솜 3개가 붙어 있는 것이 보이는가? 고리형 mRNA(Circular mRNA)에 리보솜이 여러개 결합한 것을 폴리솜(Polysome)이라 부른다.
전사가 진행됨에 따라 동시에 일어나는 mRNA의 가공을 다시 한 번 정리하면 아래의 모식도와 같다.

전사 종결에 대해선 짧게 지나가자.
3' 말단 폴리아데닐화가 일어나기 위해 mRNA가 잘려나갔다고 해서 전사가 끝나는 것이 아니다. Pol ll는 계속해서 RNA를 전사한다.
전사의 종결에 대해선 두 종류의 모델이 나와있다. 하나는 어뢰 모델(Torpedo model)이며, 하나는 다른자리 입체성 모델(Allosteric model)인데 주로 지지되는 것은 어뢰 모델이다.
어뢰 모델은 특수한 RNA 가수분해효소(RNAse)에 의해 매개되는 전사 종결 기작인데 대략 다음 모식도와 같다.

mRNA의 전사가 끝나고 난 후 생성되는 RNA는 5' 말단에 캡이 형성되어 있지 않다. Pol ll에 결합하고 있던 RNA 가수분해효소는 캡이 없는 5' 말단을 인식하여 분해를 시작한다.
(쥐의 경우 Rat1, 인간은 Xrn1이라 이름붙은 효소다)
RNA 가수분해효소는 굉장히 빠르게 RNA를 분해하지만 전에 언급했다시피 RNA 중합효소는 그다지 진행성이 좋지 못하다. RNA가 분해되는 속도보다 전사하는 속도가 훨씬 느리다.
그 결과 RNA 가수분해효소에 의해 Pol ll는 물리적으로 밀려나 DNA에서 떨어지게 된다고 여겨진다. 확실하지는 않다.
그러나 RNA 중합효소는 DNA 중합효소에 비해 굉장히 쉽게 떨어져 나갈 수 있기 때문에 상당히 설득력이 있는 모델이다.
그러면 다른자리 입체성 모델은 뭔가요?
이것은 3' 말단 폴리아데닐화 신호서열을 지난 후 Pol ll가 전사를 계속하게 되면 효소의 형태 변화가 일어나 결국 종결이 일어난다는 것이다.
이에 관련된 단백질은 C 말단 꼬리에 결합해 있다가 모종의 기전(...)을 통해 효소의 형태 변화를 유발한다고는 하나 어뢰 모델에 비해 설득력은 부족한 편이다.

기본 전사 인자





















빠져나오자마자 모자 씌우고





아니 아직도 남았나요? 지금까지는 Pol ll에 의한 mRNA의 전사에 대해서 살펴보았다. 여기서는 PolⅠ과 Pol lll에 의한 전사를 다룬다.
짧게 끝낼 것이다. 짧게.
우선, Pol ll와는 달리 PolⅠ과 Pol lll는 각각 제각기 다른 프로모터를 인지한다. 중합효소의 종류가 다르니 쉽게 납득할 수 있다.
그러나 이런 경우에도 TBP(TATA binding protein)를 필요로 한다. 어? TBP는 TFllD의 서브유닛 중 하나 아닌가요? 그러면 TFllD가 결합하나요?
그것은 아니다. TBP 서브유닛을 갖는 다른 전사 인자들이 전사에 필요하다는 것이다.
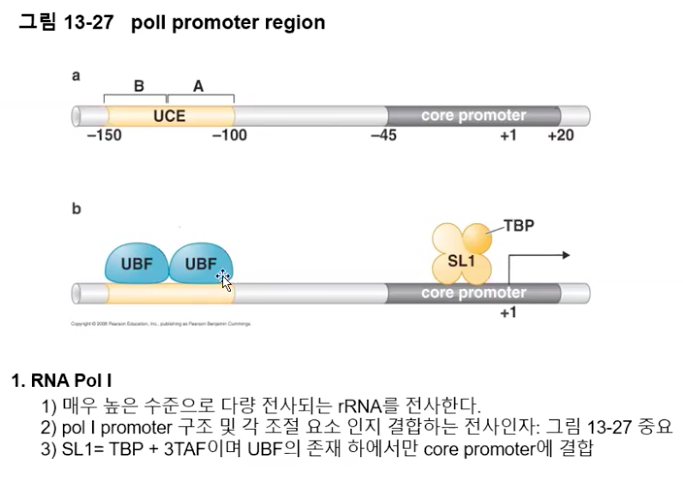
PolⅠ은 rRNA 전구체를 암호화하고 있는 단 한 종류의 유전자 발현에만 관여한다. PolⅠ에 의해 전사된 rRNA 전구체는 적절히 잘라져서 여러 rRNA 조각으로 나뉘어진다.
rRNA 유전자의 프로모터는 핵심요소(Core element)와 상류 조절요소(UCE, Upstream control element)의 두 부분으로 구성되어 있다.
사람의 경우에, rRNA 유전자의 전사 개시를 위해서 PolⅠ 외에도 SL1과 UBF라는 두 개의 다른 전사 인자가 필요하다.
이 중 TBP 서브유닛을 갖는 것이 SL1 단백질로, TBP 서브유닛과 세 종류의 TAF 서브유닛으로 구성되어 있는 단백질이다.
UBF는 상류 조절요소에 결합하여 SL1의 결합을 유도하고, SL1은 핵심요소에 결합하여 PolⅠ이 전사를 개시하게 된다.
(자세한 기전은 생략한다)
다음으로, Pol lll은 어떤 RNA를 전사하는지 기억하나? 5s rRNA을 포함한 매우 짧은 RNA들(miRNA, snRNA, siRNA 등)을 전사한다.
이들 RNA를 암호화하는 염기서열은 다양한 것들이 있는데, 따라서 Pol lll의 프로모터는 PolⅠ보다 여러 형태를 갖는다.
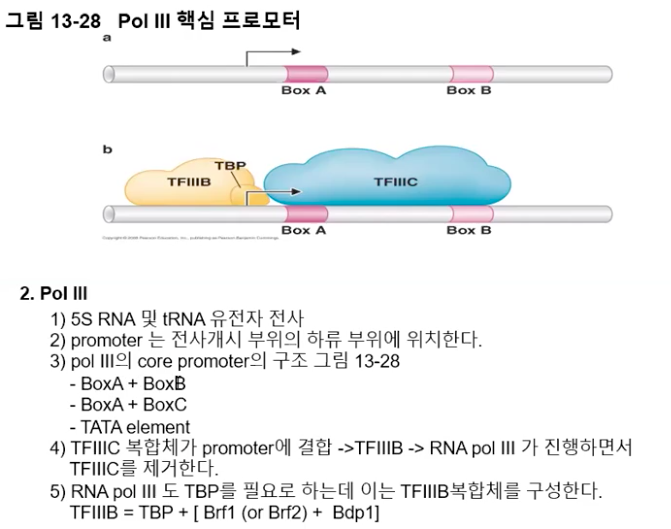
하지만 특징적인 것은, 많은 수의 Pol lll 프로모터가 전사개시점보다 아래쪽. 즉, 하류에 위치한다는 점이다.
일부 프로모터를 예로 들면, 효모 tRNA 유전자의 프로모터의 경우 상자 A와 상자 B라는 두 하류 서열을 갖고, 5s rRNA 유전자의 프로모터는 상자 A와 상자 C로 구성되어 있다.
재미있게도, Pol ll의 프로모터와 같이 TATA 박스를 갖는 유전자도 존재한다. 요는, 프로모터의 종류가 다양하다는 거다.
Pol lll에서도 마찬가지로 몇몇 전사 인자들이 필요한데, tRNA 유전자의 전사를 위해서는 TFlllB와 TFlllC라 부르는 전사인자가 관여한다.
5s rRNA의 전사를 위해선 저 두 전사인자와 함께 TFlllA도 관여한다.
TBP 서브유닛은 TFlllB가 갖고 있는데, 우선 프로모터 부위에 TFlllC가 결합하여 TFlllB의 프로모터 상류지역 결합을 유도한다.
TFⅢB가 전사 개시 부위의 상류 DNA에 결합하게 되면, 전사 개시점에 Pol lll이 결합하여 전사를 시작하게 되며, 당연히 전사가 진행되는 와중에 하류에 위치한 TFlllC는 떨어져 나가게 된다.
이것 역시 자세한 기전은 생략하도록 하겠다. 물론 rRNA와 tRNA에 대해서는 나중에 더 다룰 일이 있겠지만, 여기서는 그냥 전사의 조절이 어떻게 일어나는가. 정도만 살펴보는 것이 목적이다.
'🧬 Bio > 분자생물학' 카테고리의 다른 글
| 분자생물학 (14) - Alternative splicing, Exon shuffling, RNA editing (0) | 2020.11.30 |
|---|---|
| 분자생물학 (14) - RNA splicing (0) | 2020.11.27 |
| 분자생물학 (13) - 전사기작 (1) | 2020.11.25 |
| 분자생물학 10장 (2) - DNA 손상 (0) | 2020.10.15 |
| 분자생물학 10장 (1) - 돌연변이와 mismatch repair (1) | 2020.10.11 |